
Download

Original Article
Boosting the nutritional value of edible dahlia flowers with nano-zinc oxide (ZnO)
Orlanda Tanahiri García-González1, Oscar Cruz-Alvarez1, Yolanda Leticia Fernández-Pavía2, Oscar Sariñana-Aldaco3, María Noemí Frías-Moreno1, María del Rosario García-Mateos4, María Teresa Martínez-Damián4*, Damaris Leopoldina Ojeda-Barrios1*
1Facultad de Ciencias Agrotecnológicas, Universidad Autónoma de Chihuahua, Chihuahua, México;
2Recursos Genéticos y Productividad, Colegio de Postgraduados, Texcoco de Mora, Estado de México, México;
3Programa de Posdoctorado de la Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI), Universidad Autónoma Agraria Antonio Narro, Coahuila de Zaragoza, México;
4Departamento de Fitotecnia, Universidad Autónoma Chapingo, Texcoco de Mora, Estado de México, México
Abstract
Edible flowers of dahlia represent a novel source of functional foods, and their nutritional enhancement through agronomic zinc biofortification is scarcely explored. This study establishes, for the first time, the efficacy of -foliar-applied zinc oxide nanoparticles (ZnO NPs) as a novel biofortification strategy for this crop. The study compared foliar applications of ZnO NPs with those of zinc sulphate (ZnSO4) at 50 mg L–1 or 100 mg L–1 in dahlia flowers cv. ‘Yaretzi’. Plants were grown in open-field pots. Levels of bioactive compounds, antioxidant activity, and mineral composition were measured. Results show that ZnO NPs treatment significantly increased chroma by 20% (14.63 ± 0.87), anthocyanins by 40% (0.21 ± 0.02 mg C3G 100 g–1) and Zn content by 37% (49.7 ± 0.57 mg kg–1), compared to the treatment with ZnSO4. Antioxidant capacity (152.61 ± 2.54 µmol Fe2+/g) and protein (5.6 ± 0.03%) and fibre (30.3 ± 0.4%) were also increased. Findings indicate that foliar applications of ZnO NPs exhibit enhanced effectiveness, compared to ZnSO4, for increasing the nutrient-richness of dahlia flowers. Consequently, foliar application of ZnO NPs emerges as a promising agronomic strategy to cultivate dahlia flowers as a functional food for combating zinc deficiencies, subject to future comprehensive assessment of nanoparticle toxicology in food chain.
Key words: bioactive compounds, biofortification, functional food, mineral composition, ZnO nanoparticles (ZnO NPs)
*Corresponding Authors: Damaris Leopoldina Ojeda-Barrios, Facultad de Ciencias Agrotecnológicas, Universidad Autónoma de Chihuahua, Calle Escorza 900, Colonia Centro, CP 31000, Chihuahua, México. Email: dojeda@uach.mx; María Teresa Martínez-Damián, Departamento de Fitotecnia, Universidad Autónoma Chapingo, Carretera México-Texcoco Km. 38.5, CP 56230, Estado de México, México. Email: mmartinezda@chapingo.mx
Academic Editor: Ismail Eş, PhD, Institute of Biomedical Engineering, Old Road Campus Research Building, University of Oxford, Headington, Oxford, UK
Received: 20 August 2025; Accepted: 25 April 2026; Published: 1 June 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
The Dahlia genus (Asteraceae, Coreopsideae) is endemic to Mexico, where 36 of the world’s 42 species are discovered. The genus comprises perennial herbs and shrubs primarily adapted to terrestrial environments (American Dahlia Society [ADS], 2025; Carrasco-Ortiz et al., 2019; Sánchez-Chávez et al., 2026). Its remarkable floral diversity has established its primary value as an ornamental resource; yet, as an endemic genetic resource, it also holds significant cultural, economic, and largely untapped culinary potential (Mejía-Muñoz et al., 2020; Nsabimana and Jiang, 2011; Reyes-Santiago et al., 2024). Paradoxically, while extensive hybridization has yielded approximately 61,000 cultivars, these are derived from just four species (D. coccinea, D. pinnata, D. merckii, and D. imperialis) and represent only ~10% of the genus’s genetic diversity (Mera-Ovando and Bye-Boettler, 2006; Wang et al., 2024). This limited genetic base exists besides severe threats of extinction because of climate change, habitat alteration, and wildfire (Mejía-Muñoz et al., 2015). Scientific research now validates much of the ancestral knowledge of dahlia’s health benefits, and positions these flowers as a promising functional food. Beyond their traditional use as garnishes and their value in modern gastronomy, dahlia flowers contain a rich profile of bioactive compounds—including phenols, flavonoids and anthocyanins (Kumari et al., 2017). All these have antioxidant activity that underpins their health-promoting properties (Hernández-Epigmenio et al., 2022; Mulík et al., 2024; Mulík and Ozuna, 2024; Najar et al., 2024).
The rising global demand for functional foods is fuelled by shifting consumer preferences towards novel and health-promoting ingredients. This has led to a significant increase in interest in edible flowers (EFs). Among EFs, dahlia flowers have attracted particular attention because of their historical culinary use and rich phytochemical profile. This has positioned them as a strong candidate for dietary diversification and targeted nutritional enhancement (Costa et al., 2022; Granados-Balbuena et al., 2023; Whitley, 1985). The high antioxidant and therapeutic value of consuming either wild or cultivated dahlia flowers is receiving growing interest (Chetia et al., 2025). Recent studies have characterised their phytochemical profiles and mineral contents, and this has further validated their dietary potential (Granados-Balbuena et al., 2024a; Marcos-Gómez et al., 2025; Rivera-Espejel et al., 2019). However, critical knowledge gaps persist, particularly in: (a) biofortification strategies for EFs (an underexplored area in dahlias); (b) optimisation of agronomic management, and (c) post-harvest processing and food safety protocols. This presents a critical opportunity to expand our utilisation of the remaining 90% of dahlia species for both ornamental purposes and as innovative candidates for commercial production of EFs.
The pursuit of sustainable development and food security has emerged as a global priority, particularly in addressing persistent hunger—a challenge for many human populations globally (Cakmak and Kutman, 2017; White and Broadley, 2009). A critical aspect of this challenge is micronutrient malnutrition—the so-called ‘hidden -hunger’. Micronutrient malnutrition often stems from diets dominated by energy-dense but nutrient-poor foods as well as from physiological stressors, such as pregnancy or underlying health conditions (Rizvi et al., 2023). One of these malnutrition conditions is due to zinc (Zn) deficiency. This impairs physical development and increases susceptibility to diarrheal diseases, respiratory infections, and childhood mortality (Sharma et al., 2025). These conditions disproportionately impact marginalised communities, especially those inhabiting regions with nutrient-depleted soils. These are often characterised by high salinity or carbonate dominance, or poor physical structure (Minnocci et al., 2018). Among the strategies to minimise this deficiency is biofortification, which is the process by which the nutrient density of food crops is increased through conventional plant breeding, improved agronomic practices, and/or modern biotechnology without sacrificing any characteristics that are preferred by consumers or most importantly by farmers (Azeem et al., 2025; Sharma et al., 2025). Biofortification to create functional super-foods is gaining prominence as a scalable solution in these areas by enhancing the mineral concentrations of various edible crops by employing novel phytogenetic, genetic, or agronomic approaches (Cakmak and Kutman, 2017). Of these, agronomic biofortification offers the most immediate, expedient, and cost-effective implementation pathway. This can be achieved by working with both conventional and underutilised crops, such as dahlia (Du et al., 2019).
The effectiveness of soil applications of these mineral nutrients is influenced by multiple factors, such as soil water content, irrigation practices, soil physicochemical properties (pH, texture, structure, etc.), nutrient solubility, chemical source, and dose (Hatamian et al., 2018). Given these limitations, foliar sprays emerge as a viable alternative to soil applications, as they increase the precision of application and also offer advantages in terms of efficiency, speed, adaptability, and immediate correction of deficiencies (Davarpanah et al., 2017). Furthermore, by requiring smaller amounts of fertiliser, foliar sprays reduce the ecological impact on agroecosystems compared to soil application (Niu et al., 2021). The use of Zn oxide nanoparticles (ZnO NPs) has gained relevance as an inorganic source of Zn with a stable chemical structure, high specific surface area, and redox capacity (Bala et al., 2019; Karimian and Samiei, 2023). These characteristics provide ZnO NPs greater penetration into leaf tissues and mobility within the plant while also minimising losses because of leaching or immobilisation in soil—these two factors being -typical drawbacks with conventional fertilisers, such as zinc -sulphate (ZnSO4), zinc nitrate (Zn(NO3)2) or Zn chelates (ethylenediamine-N,N'-bis(2-hydroxyphenylacetic acid [EDDHA], ethylenediaminetetraacetic acid [EDTA], and diethylenetriaminepentaacetic acid [DTPA]) (Bautista-Diaz et al., 2021; Mahdavi et al., 2022). After foliar application, ZnO NPs appear to enter the plant mainly through the stomata, they accumulate initially in the apoplast and then dissolve progressively, releasing Zn2+ absorbed by the mesophyll cells, while part of a nanoparticulate fraction may also be internalised. This may account for their distinct distribution within leaf tissues, compared to ZnSO4 (Zhu et al., 2020). A gradual release of Zn may improve Zn accumulation and translocation and support Zn-dependent metabolic processes, including Cu/Zn-superoxide dismutase activity, thereby contributing to improved growth and stress tolerance (Rameshraddy et al., 2017).
Despite the promising agronomic benefits of ZnO NPs, their application in edible crops necessitates careful toxicological assessment (Hou et al., 2018). Recent studies have highlighted that nanoparticle size critically determines respiratory deposition, with a 67.5% reduction in deposition fraction as particle size increases from 10 nm to 100 nm (Khadanga and Mishra, 2024). The -multiple-path particle dosimetry modelling confirms significant nanoparticle retention in the pulmonary lobes, while in vivo studies demonstrate that inhaled nanoparticles can translocate to the bloodstream and brain, potentially triggering oxidative stress, inflammation, and tissue damage. Additional concerns include dermal absorption, environmental persistence, and limited knowledge about long-term health consequences (Kah et al., 2021). Consequently, while this study focuses on establishing biofortification efficacy, comprehensive risk evaluations, including respiratory exposure assessment, translocation studies and dietary safety analysis, remain essential before commercial implementation in the food chain.
Building upon these advantages, this study pioneers the Zn biofortification of edible dahlia flowers using foliar-applied nanoparticles. This innovative approach significantly enhances Zn accumulation and bioactive compound content, establishing a new paradigm for nutritional improvement in ornamental-edible species. Furthermore, ZnO NPs are compatible with sustainable management systems, as their use reduces both dose and application frequency needed to fulfil the crop’s nutritional demands (Almendros et al., 2022). This study, therefore, evaluates the effects of foliar applications of ZnO NPs versus ZnSO4 on Zn enrichment, bioactive compounds, and mineral composition of dahlia cv. ‘Yaretzi.’ It is anticipated that the findings will provide a technical foundation for leveraging this underutilised genetic resource, supporting the plant’s culinary diversification while maximising its nutritional potential.
Materials and Methods
Experimental site and plant material
The study was conducted from March 2024 to November 2024 under open-field conditions in the horticulture area adjacent to the floriculture unit at the Autonomous University Chapingo (UACh) located in Texcoco de Mora, State of Mexico, Mexico (19°29’23” N, 98°52’24” W). The site lies at an elevation of 2,268 m above sea level, with an average annual temperature of 15.6°C. Dahlia plants (Dahlia × hortorum) cv. ‘Yaretzi’ were propagated from tuberous roots collected from the ‘San Martín’ experimental field (19°29’58” N, 98°52’44” W) managed by the Department of Plant Science (UACh). Plant propagation began in late March 2024 using five 210-L high-density polyethylene containers (≈40 tuberous roots/container) filled with a 2:1 (v/v) peat–perlite substrate. Sprout emergence occurred 15 days after planting (DAP), with subsequent transplantation at 40 DAP into individual 6-L black polystyrene bags containing the same substrate mixture (Figures 1A and 1B). Drip irrigation was administered at 0.5–3.0 L/plant per session (1–2 times weekly), with volume and frequency adjusted according to the prevailing weather conditions (temperature and relative humidity) and crop phenological stage. Nutrient management followed a 120–0–200 (N–P2O5–K2O) fertilisation regimen, split into two applications: first at transplantation, and the second just pre-flowering (BBCH stage 51). Throughout the trial, integrated pest management and weed control followed established commercial protocols for ornamental dahlia production.
Figure 1. Plant material of dahlia cv. ‘Yaretzi’. (A) General view of the crop under open-field conditions at the experimental site; (B) a representative in florescence exhibiting characteristic morphology of the cultivar.
Experimental design and sampling
The study employed a completely randomised design with five treatments and four biological replicates per treatment (n = 4). Each biological replicate constituted an experimental unit of three plants (60 plants in total). Five treatments were evaluated: (T1) absolute control (no Zn application); (T2 and T3) two concentrations of ZnO nanoparticles (50 mg L–1 and 100 mg L–1; Investigación y Desarrollo de Nanomateriales S.A. de C.V., Mexico); and (T4 and T5) two concentrations of ZnSO4 (50 mg L–1 and 100 mg L–1; Merck®, Germany). All Zn treatments included 100 μL L–1 of Silwet® L-77 adjuvant (trisiloxane-based; Arista-Lifescience, Mexico). Foliar applications were made at three phenological stages: 3 and 5 weeks post-transplanting (vegetative phase) and at flower bud stage (pre--anthesis). Applications were made between 08:00 and 09:00 h using a 0.5-L fine sprayer to ensure complete leaf coverage (both surfaces), with the substrate protected by a plastic film to prevent soil contamination. Inflorescences were asynchronously harvested beginning 25 August 2024 using sterilised pruning shears. Samples were collected at the precise phenological stage when the stamens became visible in the first row of tubular florets. To minimise post-harvest stress, harvesting was conducted during optimal morning hours (between 07:00 and 09:00 h). Flower heads were immediately transported for processing in dry ice-cooled containers to the Plant Physiology Laboratory (Department of Plant Science, UACh).
At the laboratory, the capitula were separated manually into individual florets and subjected to lyophilisation using a BK-F10PT freeze dryer (BIOBASE®, USA). For each replicate (n = 4 per treatment), 50-g samples of ray florets (ligules) were evenly distributed in standard trays (180 × 20 mm) and processed at -56°C and 0.1 mbar for 72 h until constant weight was achieved. The freeze dryer was pre-stabilised to target parameters before each run to ensure consistent processing conditions. The resulting lyophilised material was then crunched into a fine powder using a mortar and pestle ready for subsequent analysis of bioactive compounds (total phenols [TP], total flavonoids [TFl], and total anthocyanins [TAn]) and antioxidant capacity (AC).
Reagents
All chemicals used in this study, including those for the determination of photosynthetic pigments, sucrose, bioactive compounds, antioxidant capacity, and mineral composition, were of analytical grade and purchased from Sigma-Aldrich, USA.
Characterisation of nanoparticles
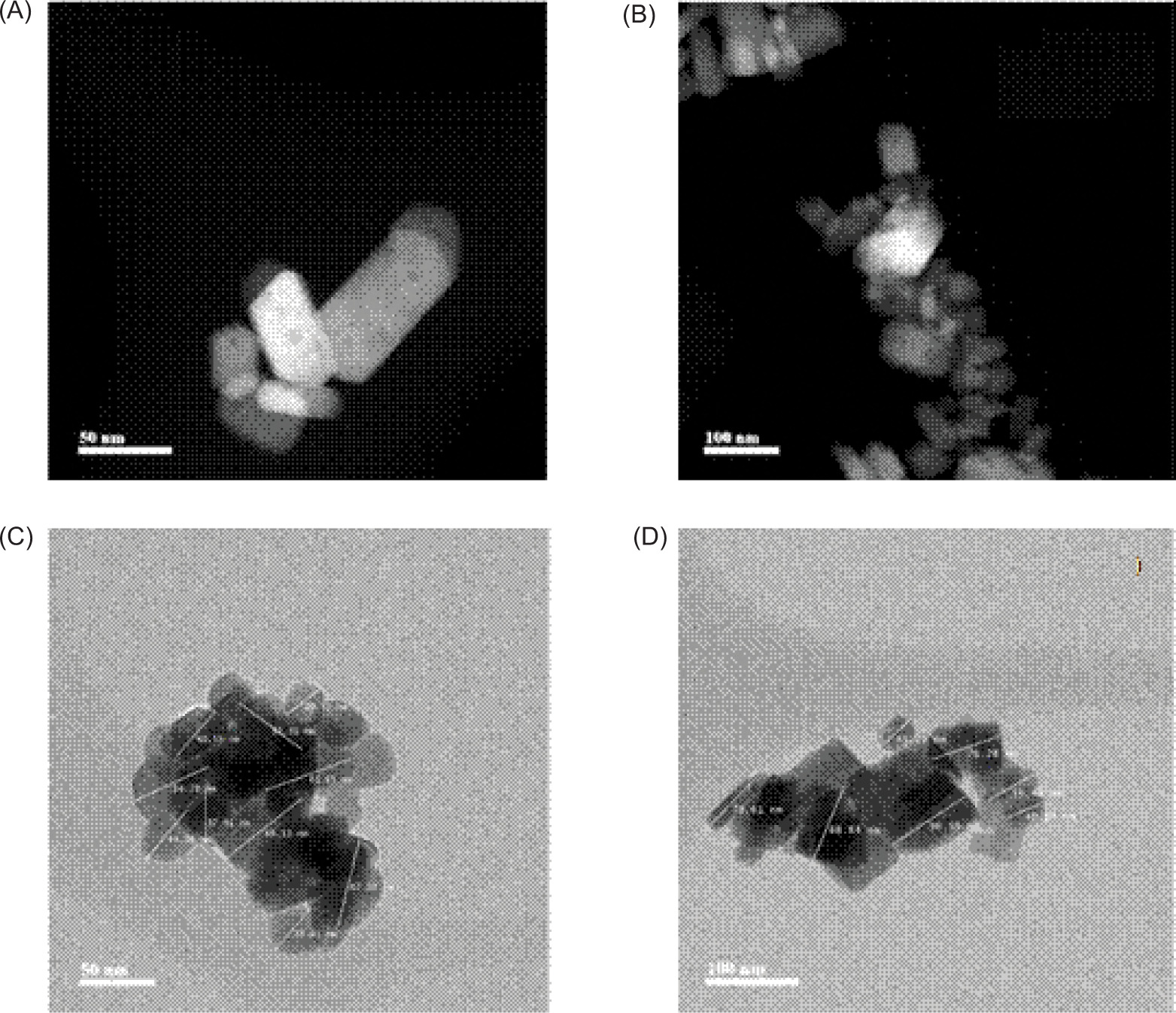
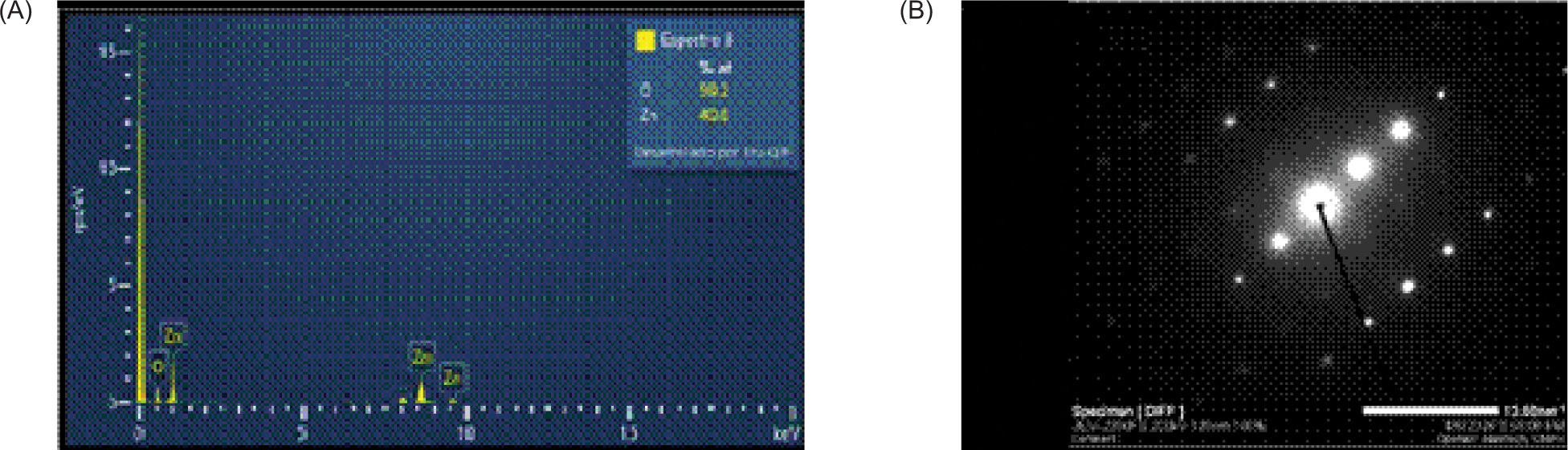
Zinc oxide powder was applied as a nanofertiliser. For characterisation, the sample was dispersed by ultrasonication and deposited onto a 300-mesh copper grid coated with a Lacey Formvar/carbon support film (Ted Pella, USA). The analysis was carried out using a JEOL JEM-2200FS-CS transmission electron microscope (JEOL, Japan) operated at 200 kV. Morphology was examined by scanning transmission electron microscopy (STEM), crystallinity by selected-area electron diffraction (SAED), and chemical composition and elemental mapping by energy-dispersive X-ray spectroscopy (EDS) using an Oxford UltimR-Max detector (Oxford Instruments, UK). The material exhibited nanometric dimensions, with particle sizes ranging from 29.52 nm to 116.80 nm, with an average size of 64.66 nm (Figures 2C and 2D). The particles were predominantly elongated, with well-defined faces resembling small prisms or rods, and some agglomeration was observed (Figures 2A and 2b). Diffraction patterns indicated a highly ordered crystalline material (Figure 3B). EDS spectra obtained at different points showed only Zn and O signals, with no evident impurities detected within the limits of the technique, suggesting a relatively homogeneous composition (Figure 3A).
Figure 2. Morphological and dimensional characterisation of ZnO nanomaterial applied as a nanofertilizer. (A and B): -nanoparticle morphology obtained by STEM; (C and D): particle size measurements obtained by transmission electron -microscopy (TEM).
Figure 3. ZnO nanoparticles: (A) elemental analysis (chemical composition) by energy-dispersive X-ray spectroscopy (EDS), and (B) crystalline structure by selected-area electron diffraction (SAED).
Colour, total soluble solids (TSS), and titratable acidity (TA)
Flower colour parameters were quantified using a portable SP62 sphere colorimeter (X-Rite®, USA), measuring CIELAB coordinates: L* (lightness), C* (chroma), and h° (hue angle). For physicochemical analyses, TA was determined following the method described by Tomasik (2003): briefly, a 0.5-g sample was homogenised with 10-mL distilled water using an Ultra-Turrax T25 homogeniser (IKA®, USA) for 2 min, filtered, then titrated against 0.1-N NaOH with 2% phenolphthalein as indicator (results expressed as %citric acid). TSS were measured by placing filtrate droplets on a PAL-1 digital refractometer prism (ATAGO®, Japan), with results reported in °Brix.
Photosynthetic pigment and leaf area
The extraction and quantification of chlorophylls a and b (Chl a and Chl b) and total carotenoids (TC) was carried out by the method described by Wellburn (1994). Briefly, a sample of fresh leaf tissue (5 g) was placed in a 100-mL flask with 80 mL of 80% acetone. Absorbance was recorded at 665 nm, 653 nm, and 470 nm using an ultraviolet (UV)/visible spectrophotometer Lambda 25® (PerkinElmer, USA) with 1-cm pathlength quartz cuvettes. Pigment concentrations were calculated using the following equations:
Chl a = 12.72A655 - 2.59A653,
Chl b = 20.36A653 - 5.19A665,
TC = (1,000A470 - 1.63 Chl a - 104.96 Chl b) ÷ 221.
To quantify foliar development, leaf area (LA) was determined according to a procedure described by Liu et al. (2022) with slight modifications. Briefly, prior to flower harvest, all leaves were measured using a CI-202 Laser Leaf Area Meter (CID Bio-Science, USA). The results are expressed in cm2.
Sucrose
The sucrose was quantified according to the method described by Irigoyen et al. (1992) with slight modifications: Briefly, 0.5-g fresh dahlia ligulate florets was homogenised three times, the first time with 5 mL of 96% ethanol and then twice with 5 mL of 70% ethanol. The homogenate was centrifuged at 3,740×g for 10 min at 4°C. For colorimetric analysis, 0.1 mL of supernatant was reacted with 3 mL of freshly prepared anthrone reagent (1% w/v anthrone in concentrated H2SO4). The mixture was incubated in a boiling water bath (100°C) for exactly 10 min, then immediately cooled on ice. Absorbance was measured at 650 nm against a reagent blank using a Lambda X® spectrophotometer (PerkinElmer) with 1-cm pathlength quartz cuvettes. Concentration of sucrose was determined from a -standard curve (0–100 μg mL sucrose) and expressed as mg g-1 fresh weight (FW).
Bioactive compounds
Extraction
The extraction of bioactive compounds was conducted by the method described by Aguiñiga-Sánchez et al. (2017) with modifications. Four biological replicates (n = 4) were analysed per treatment. Lyophilised floral tissue (0.50 ± 0.02 g per replicate) was homogenised in 5 mL of 80% methanol (v/v, high performance liquid chromatography [HPLC] grade) using a IKA T25 Ultra-Turrax at 10,000 rpm for 2 min. The homogenate was centrifuged at 6,000×g for 10 min at 4°C. The supernatant was used to quantify phenols, flavonoids, anthocyanins, and antioxidant capacity as described below. For variables quantified from lyophilised tissue, results were converted on an FW basis using the moisture content determined for each treatment according to the following equation:
Value on FW basis = value on DW basis × (100 – -moisture, %) ÷ 100.
The corresponding moisture values used for conversion were obtained from the proximate analysis of the same treatment.
Phenols, flavonoids, total anthocyanins, and ascorbic acid (AA)
Total phenols (TP) were quantified according to the Folin–Ciocalteu method described by Silveira et al. (2020). Two 0.250-mL samples of the above supernatant were chosen, to which were added 0.750 mL of 2% (v:v) Na2CO3 and 0.250 mL of 50% Folin–Ciocalteau. The mixture was incubated for 1 h and the absorbance was measured at 725 nm on a Lambda 25® UV-visible spectrophotometer (PerkinElmer). Quantification was performed using a gallic acid standard curve (0–500 mg L–1, R2 > 0.99), with results expressed as mg gallic acid equivalent (GAE) per 100 g FW (mg GAE 100 g–1 FW).
Total flavonoids were determined according to the method described by Chang et al. (2002). Briefly, 0.250 mL of the above supernatant was mixed with 0.075 mL of 5% (w/v) sodium nitrite (NaNO2) and vortexed for 5 min. Subsequently, 0.150 mL of 10% (w/v) aluminium chloride (AlCl3) was added, followed by 0.500 mL of 1-M sodium hydroxide (NaOH) and 2.025 mL of deionised water. The reaction mixture was incubated for 40 min at ambient temperature (25 ± 2°C) in the dark. Absorbance was measured at 510 nm using a Lambda 25® UV-Vis spectrophotometer (PerkinElmer). Quantification was performed against a quercetin standard curve (0–200 mg L–1, R2 > 0.98), with results expressed as mg quercetin equivalent (QE) per 100 g FW (mg QE 100 g–1 FW).
Total anthocyanin content was quantified using the pH differential method (Giusti and Wrolstad, 2001), with modifications. Two 0.5-mL aliquots of the above supernatant were separately mixed with either 2.0 mL of 0.025-M potassium chloride buffer (pH 1.0) or 2.0 mL of 0.4-M sodium acetate buffer (pH 4.5), followed by vortexing for 1 min. Absorbance measurements were accomplished at two wavelengths: 520 nm to quantify anthocyanins at their peak absorption, and 700 nm to correct for potential light scattering from sample turbidity, using a Lambda 25® UV-Vis spectrophotometer (PerkinElmer). Anthocyanin concentration was calculated by comparing the difference in absorbance between two pH conditions, with results expressed as milligram of cyanidin-3-glucoside (C3G) equivalent per 100 g of FW (mg C3G100 g-1 FW). This calculation incorporated molar absorptivity (26,900 L•cm–1•mol–1) and molecular weight (449.2 g/mol) of C3G as reference standards.
Antioxidant capacity
The antioxidant capacity of flower extracts was comprehensively evaluated using three validated assays, each performed with four biological replicates (n = 4) and three technical replicates per sample. The methods were: ABTS 2,2’-azino-bis(3-ethylbenzothiazoline-6--sulfonic acid) radical scavenging (ABTS), ferric reducing antioxidant power (FRAP), and 2,2-diphenyl-1-picrylhydrazyl (DPPH). The ABTS assay was carried out as described by Re et al. (1999), for which ABTS solution was prepared by mixing equal volumes of 7 mM of ABTS reagent ((2,2’-azino-bis(3-ethylbenzothiazoline)-6-sulfonic acid)) and potassium persulfate (K2S2O8) at 2.45 mM (w:v), which was left to stand for 116 h at room temperature. Subsequently, 20% ethanol was diluted to obtain an absorbance value of between 0.24 and 0.67 at 734 nm; then, 10 µL of the sample diluted in ethanol + 90 µL of distilled water + 3 mL of ABTS applied every 30 s were added and left to react for 15 min. The absorbance was obtained at 734 nm and the results are expressed as µmol TE (Trolox equivalent) g-1.
The FRAP assay was used to determine antioxidant capacity as described by Benzie and Strain (1996). Briefly, 2.3 mL of FRAP reagent was mixed with 0.7 mL of aqueous extracts. The mixture was then incubated at 37°C for 30 min in the dark. This mixture was then filtered with Whatman® 40 qualitative filter paper (Sigma-Aldrich) and centrifuged at 11,500 rpm for 20 min. The absorbance value was obtained at 593 nm and the results were expressed as µmol Fe2+ g-1. The DPPH assay was carried out as described by Brand-Williams et al. (1995) with modifications. Briefly, 1 mL of 0.1-mM solution of DPPH reagent in methanol was mixed with 2 mL of extract (v/v) and the mixture allowed to stand for 15 min at room temperature before obtaining the absorbance value at 517 nm, where 20% (v:v) methanol was used as a blank. Antioxidant capacity was expressed on a dry weight (DW) basis, without conversion to FW, and DPPH values are presented as %inhibition.
Ascorbic acid
The ascorbic acid content was determined using the 2,6-dichlorophenol-indophenol (DCPI) titration method (Association of Official Analytical Chemists [AOAC] Official Method 967.21, Tillmans, 1990). Fresh plant tissue (3.0 ± 0.1 g) was homogenised in 50 mL of 2% (w/v) oxalic acid, which served as both extraction solvent and stabilising agent (preventing ascorbic acid oxidation at pH < 3.0). A 10-mL aliquot of filtered homogenate was titrated against standardised DCPI solution (Tillmans reagent) until a persistent pink endpoint was obtained (15–30 s). Acid ascorbic content was calculated from a freshly prepared ascorbic acid standard curve (0–100 mg L–1, R2 ≥ 0.995) and expressed as milligram ascorbic acid per 100 g FW (mg AA 100 g–1 FW). All extractions and titrations were performed under dim light to minimise photo-oxidation.
Proximate composition
The proximate composition of flower samples was determined by following standardised methods (Tomasik, 2003) with four biological replicates (n = 4) per treatment. The results were expressed as percentage values. Total energy of one serving (100 g FW) was calculated according to the following equation:
Total energy = (Energy content of 1 g protein × g protein of sample) + (Energy content of 1 g fat × g fat of sample) + (Energy content of 1 g carbohydrate × g carbohydrate of sample) (Chahdoura et al., 2015).
Mineral nutrients in flowers
The flowers were harvested and transported for analysis to the Fruit Tree Physiology Laboratory at Universidad Autonoma Chapingo, Mexico, where extraction and quantification of mineral nutrients was carried out by using the method described by Ontiveros-Capurata et al. (2022). Briefly, flowers were triple washed (1) with tap water, then (2) with 4-N HCl, and lastly (3) with deionised water. Surface water was allowed to evaporate completely at room temperature and leaves were dried at 75°C for 24 h in a Heratherm VCA 230® oven (Thermo Scientific™, Waltham, USA). Each sample was homogenised in a Willey R-TE-650/1 mill with 1-mm mesh (Tecnal, Brazil). The extraction and quantification of total-N were determined by the Kjeldhal method (Novatech®, USA and Micro Kjeldahl Labconco®, USA). Total P was determined by using the ammonium metavanadate method (NH4VO3) (Thermo Scientific™). The extraction of K+, Ca2+, Mg2+, Fe2+, Mn2+, Cu2+, Zn2+, and Na+ was carried out by triacid digestion (HNO3, HClO4, and H2SO4) (25 mL of mixture in a 10:10:25 ratio) using 25 mL of acid mixture on a hot plate under a fume-hood. Analyte quantifications was carried out using an Analyst 100® atomic absorption spectrophotometer (PerkinElmer). Results are reported as g kg-1 for macronutrients and mg kg-1 for micronutrients. The B concentration was determined by a colorimetric method using azomethine H reagent for plant ash extract (Gaines and Mitchell, 1979).
Data analysis
To validate the analysis of variance (ANOVA), Bartlett’s test (homogeneity of variance) and Kolmogorov–Smirnov test (normality) were applied (P ≤ 0.05) (Sokal and Rohlf, 1995). Subsequently, a simple rank analysis of variance and a multiple comparison of means were performed using Tukey’s test (P ≤ 0.05). Results were presented as mean ± standard error of mean (SEM) for four biological replicates (n = 4). In addition, Pearson’s correlation matrix was also constructed between mineral nutrients and petal quality variables. In all procedures, the SAS software, version 9.0 (SAS Institute, Cary, NC, 2002) was used.
Results
Colour, total solid soluble, and titratable acidity
As presented in Table 1, colour parameters (L*, C*, and h°), TSS, and TA were evaluated across treatments. The ZnO NPs treatment (50 mg L–1 and 100 mg L–1) significantly increased chroma values (C* = 14.63 ± 0.87 for both concentrations; P < 0.05), compared to controls. This enhanced colour saturation represented a key quality attribute, potentially improving both pollinator attraction and consumer appeal for ornamental/culinary uses. However, lightness (L*) and hue angle (h°) values remained statistically unchanged (P > 0.05) across all treatments. Similar stability was observed in other quality markers, with no significant treatment effects on either TSS content or TA levels under experimental conditions.
Table 1. Effects of ZnO nanoparticles and ZnSO4 on colour parameters and quality of edible flowers of dahlia cv. ‘Yaretzi’.
| Treatment (mg L–1) |
Colour | Total soluble solids (°Bx) | Titratable acidity (% citric acid) | ||
|---|---|---|---|---|---|
| L | Chroma | h° | |||
| Control | 60.36 ± 1.78a | 12.49 ± 1.04b | 64.38 ± 0.72a | 0.86 ± 0.02a | 3.23 ± 0.04a |
| 50ZnO NPs | 61.83 ± 1.56a | 14.63 ± 0.87a | 65.13 ± 1.34a | 0.88 ± 0.01a | 3.13 ± 0.03a |
| 100ZnO NPs | 62.90 ± 1.86a | 14.51 ± 0.60a | 65.33 ± 0.98a | 0.89 ± 0.02a | 3.16 ± 0.04a |
| 50ZnSO4 | 59.90 ± 2.11a | 12.08 ± 0.76b | 66.10 ± 1.12a | 0.84 ± 0.02a | 3.18 ± 0.04a |
| 100ZnSO4 | 60.46 ± 1.97a | 12.65 ± 0.81b | 65.34 ± 1.23a | 0.87 ± 0.02a | 3.17 ± 0.04a |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ statistically (P ≤ 0.05; Tukey’s test).
L: lightness; h°: hue angle; ZnO NPs: zinc oxide nanoparticles; ZnSO4: zinc sulphate.
Table 2. Effects of ZnO nanoparticles and ZnSO4 on photosynthetic pigment content and leaf area in dahlia cv. ‘Yaretzi’.
| Treatment (mg L-1) | Chlorophyll (μg g-1) | Total carotenoids (µg ß-carotene g-1) |
Leaf area (cm2) | |
|---|---|---|---|---|
| a | b | |||
| Control | 1.04 ± 0.20a | 0.173 ± 0.02c | 15.96 ± 1.40a | 8.09 ± 0.002b |
| 50ZnO NPs | 0.96 ± 0.10a | 0.254 ± 0.09b | 15.92 ± 1.48a | 8.01 ± 0.001b |
| 100ZnO NPs | 0.92 ± 0.18a | 0.322 ± 0.1a | 16.01 ± 2.61a | 8.34 ± 0.001a,b |
| 50ZnSO4 | 1.02 ± 0.21a | 0.376 ± 0.08a | 16.20 ± 1.31a | 8.90 ± 0.002a |
| 100ZnSO4 | 1.1 ± 0.18a | 0.298 ± 0.11a,b | 16.19 ± 3.78a | 8.76 ± 0.001a |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ significantly (P ≤ 0.05; Tukey’s test). Data is expressed on a fresh weight (FW) basis. ZnO NPs - zinc oxide nanoparticles; ZnSO4 - zinc sulphate.
Photosynthetic pigments and leaf area
In dahlia flowers, Zn treatments had selective effects on pigment profiles. Foliar application of 100 mg L–1 ZnO NPs and ZnSO4, together with 50 mg L–1 ZnSO4, significantly increased both Chl b and TC contents. Chl b reached 0.322 ± 0.10, 0.376 ± 0.08, and 0.298 ± 0.11 μg g–1 FW, whereas TC reached 8.34 ± 0.001, 8.90 ± 0.002, and 8.76 ± 0.001 μg β-carotene equivalent g–1 FW. In contrast, no significant changes were observed in leaf area (15.92–16.20 cm2) or Chl a concentration (0.92–1.04 μg g–1 FW). These findings showed that Zn acts as a key micronutrient that preferentially modulates biosynthesis pathways of accessory pigments while maintaining structural components of photosynthetic apparatus.
Sucrose and bioactive compounds
Our results revealed a differential accumulation pattern of secondary metabolites in response to Zn treatments (Table 3). While neither ZnO NPs nor ZnSO4 significantly affected sucrose concentration (P > 0.05), other metabolites showed distinct responses: TP increased -significantly (P ≤ 0.05) with both ZnO NP treatments (6.56 ± 1.19 and 8.25 ± 1.08 mg GAE 100 g–1 FW for 50 mg L–1 and 100 mg L–1, respectively), whereas ascorbic acid content varied across all treatments (0.85 ± 0.04–1.01 ± 0.01 mg 100 g–1 FW). Total flavonoids reached their highest concentrations under 100 mg L–1 treatments (8.20 ± 1.44 and 8.21 ± 1.53 mg QE 100 g–1 FW for ZnO NPs and ZnSO4, respectively). Notably, TAn showed the most selective response, with a significant increase (0.21 ± 0.02 mg C3G 100 g–1 FW) only under 100 mg L–1 ZnO NPs, suggesting a nanoparticle-specific activation of metabolic pathways.
Table 3. Effects of zinc source (ZnO NPs vs. ZnSO4) on accumulation of sucrose and bioactive compounds in edible flowers of dahlia cv. ‘Yaretzi’.
| Treatment(mg L–1) | Sucrose (mg g–1) |
Total phenols (mg GAE 100 g–1) | Total flavonoids (mg QE 100 g–1) | Total anthocyanins (mg C3G 100 g–1) | Ascorbic acid (mg AA 100 g–1) |
|---|---|---|---|---|---|
| Control | 11.87 ± 1.2a | 6.56 ± 1.19b | 5.86 ± 1.52c | 0.15 ± 0.03b | 0.85 ± 0.04b |
| 50ZnO NPs | 11.89 ± 1.4a | 8.22 ± 1.03a | 8.20 ± 1.44a | 0.16 ± 0.03b | 1.01 ± 0.01a |
| 100ZnO NPs | 13.24 ± 1.39a | 8.25 ± 1.08a | 8.21 ± 1.53a | 0.21 ± 0.02a | 0.95 ± 0.06a |
| 50ZnSO4 | 12.98 ± 1.23a | 7.51 ± 1.04a,b | 6.15 ± 1.22c | 0.16 ± 0.03b | 0.99 ± 0.03a |
| 100ZnSO4 | 13.55 ± 1.45a | 7.72 ± 1.17a | 7.24 ± 1.08a | 0.14 ± 0.02b | 0.98 ± 0.02a |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ significantly (P ≤ 0.05; Tukey’s test). Data for total phenols, flavonoids, and anthocyanins is expressed on a fresh weight (FW) basis, converted using sample moisture content.
GAE: gallic acid equivalent; QE: quercetin equivalents; C3G: cyanidin-3-glucoside; AA: ascorbic acid; ZnO NPs: zinc oxide nanoparticles; ZnSO4: zinc sulphate.
Antioxidant capacity
Edible flowers, particularly those of dahlia cv. ‘Yaretzi’, represent a valuable dietary source of bioactive compounds with demonstrated antioxidant capacity (Granados-Balbuena et al., 2023; Ojeda-Barrios et al., 2026). Our results indicated that Zn biofortification significantly modulated antioxidant capacity, although the effects varied between analytical methods. The ABTS assay revealed increased antioxidant capacity with 100 mg L–1 ZnO NPs (73.94 ± 1.90 µmol TE/g) and both ZnSO4 concentrations (73.90 ± 2.70 and 75.03 ± 1.75 µmol TE/g for 50 mg L–1 and 100 mg L–1, respectively). More strikingly, ZnO NPs at both concentrations (50 mg L–1 and 100 mg L–1) showed maximum enhancement in ferric reducing capacity (FRAP), with values of 151.73 ± 1.79–152.61 ± 2.54 µmol Fe2+/g. These findings not only confirmed the functional potential of dahlia flowers but also highlighted the superior efficacy of ZnO NPs in boosting antioxidant properties. Particularly strong response in the FRAP assay suggested that these treatments preferentially enhanced electron transfer-based antioxidant mechanisms, underscoring the importance of formulation (nanoparticles vs. conventional salts) in biofortification strategies.
Proximate composition
The values obtained from proximate analysis are shown in Table 5. Dahlia plants treated with ZnO NPs exhibited superior nutritional profiles compared to the control group, with significant increases in several key parameters. Flowers from treated plants showed higher moisture content (91.9 ± 0.02% and 91.2 ± 0.01%), protein levels (5.2 ± 0.01% and 5.6 ± 0.03%), crude fat content (4.6 ± 0.01% and 4.6 ± 0.03%), total carbohydrates (62.1 ± 0.5% and 62.5 ± 0.6%), and total energy value per 100 g FW (1,543.0 ± 3.6 kcal and 1,565.2 ± 2.9 kcal). Notably, the application of 100 mg L–1 ZnO NPs further enhanced fibre content (30.3 ± 0.4%) and ash percentage (13.3 ± 0.06%). In contrast, crude fat calories remained statistically unchanged across all treatments, with values ranging between 40.9 ± 0.8% and 41.2%. These results demonstrated that ZnO NP biofortification could effectively improve the nutritional quality of dahlia flowers without compromising their lipid--derived energy content.
Table 4. Antioxidant capacity modulation in edible flowers of dahlia cv. ‘Yaretzi’ following ZnO NP and ZnSO4 treatments.
| Treatment(mg L–1) | ABTS(µmol TE [Trolox equivalents/g]) | FRAP(µmol Fe2+/g) | DPPH(DPPH% inhibition) |
|---|---|---|---|
| Control | 65.66 ± 1.1b | 145.23 ± 1.29b | 75 ± 0.3a |
| 50ZnO NPs | 65.56 ± 1.8b | 151.73 ± 1.79a | 74 ± 0.3a |
| 100ZnO NPs | 73.94 ± 1.9a | 152.61 ± 2.54a | 76 ± 0.3a |
| 50ZnSO4 | 73.90 ± 2.70a | 146.84 ± 1.72b | 73 ± 0.1a |
| 100ZnSO4 | 75.03 ± 1.75a | 125.02 ± 1.76b | 76 ± 0.2a |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ significantly (P ≤ 0.05; Tukey’s test). Data is expressed on a DW basis.
ABTS: 2,2'-azino-bis-3-ethylbenzthiazoline-6-sulfonic acid; FRAP: ferric reducing/antioxidant power; DPPH: 1,1-diphenyl-2-picrylhydrazyl; ZnO NPs: zinc oxide nanoparticles; ZnSO4: zinc sulphate.
Table 5. Nutritional composition (proximate analysis) of edible dahlia flowers (cv. ‘Yaretzi’) biofortified with ZnO NPs or ZnSO4.
| Treatment (mg L–1) | Moisture (%) | Crude protein (%) | Crude fat (%) | Fibre (%) |
|---|---|---|---|---|
| Control | 89.3 ± 0.03b | 4.6 ± 0.02c | 4.5 ± 0.02b | 28.3 ± 0.3c |
| 50ZnO NPs | 91.9 ± 0.02a | 5.2 ± 0.01a,b | 4.6 ± 0.01a | 29.3 ± 0.5b |
| 100ZnO NPs | 91.2 ± 0.01a | 5.6 ± 0.03a | 4.6 ± 0.03a | 30.3 ± 0.4a |
| 50ZnSO4 | 89.7 ± 0.01b | 4.8 ± 0.01b,c | 4.6 ± 0.03a | 28.5 ± 0.4b,c |
| 100ZnSO4 | 89.4 ± 0.02b | 4.9 ± 0.05b | 4.6 ± 0.05a | 28.9 ± 0.3b |
| Ash (%) | Total carbohydrates (%) | kcal/100 g) | Calories of fat | |
| Control | 12.9 ± 0.02c | 60.0 ± 0.4c | 1489.9 ± 2.9c | 40.9 ± 0.8a |
| 50ZnO NPs | 12.9 ± 0.03c | 62.1 ± 0.5a | 1543.0 ± 3.6a | 41.2 ± 1.2a |
| 100ZnO NPs | 13.3 ± 0.06a | 62.5 ± 0.6a | 1565.2 ± 2.9a | 41.2 ± 1.2a |
| 50ZnSO4 | 12.8 ± 0.04d | 60.1 ± 0.6c | 1497.0 ± 3.8c | 41.2 ± 1.4a |
| 100ZnSO4 | 13.0 ± 0.02b | 61.0 ± 0.4b | 1517.1 ± 3.9b | 41.2 ± 1.3a |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ significantly (P ≤ 0.05; Tukey’s test). Data is expressed on a DW basis.
TEOS: total energy per 100 g of fresh weight (FW); ZnO NPs: zinc oxide nanoparticles; ZnSO4: zinc sulphate.
Mineral nutrients in flowers
Table 6 presents the mineral profile of zinc-biofortified dahlia flowers cultivated under different Zn treatments. Among the macronutrients analysed and treatments applied, plants treated with a high dose of ZnO NPs showed increased concentrations of P (3.1 ± 0.06 g kg–1), K+ (12.9 ± 1.30 g kg–1), Mg2+ (2.7 ± 0.06 g kg–1), and Ca2+ (2.2 ± 0.05 g kg–1), with the exception of total N (20.3 ± 0.58 g kg–1 and 21.3 ± 0.78 g kg–1 for low and high doses, respectively). Notably, the 50-mg L–1 ZnO NP treatment showed statistically similar results for Ca2+ accumulation (2.2 ± 0.05 g kg–1). The ZnO NP treatments induced distinct patterns of mineral accumulation in dahlia flowers. The 50-mg L–1 ZnO NP dose significantly enhanced iron (63.2 ± 3.73 mg kg–1) and copper (12.8 ± 0.61 mg kg–1) contents, while the 100-mg L–1 concentration proved more effective for Zn (39.7 ± 0.57 mg kg–1) and Na (15.8 ± 0.1 mg kg–1) accumulations. Similarly, both ZnO NP concentrations improved boron uptake (23.3 ± 2.20 and 21.9 ± 3.11 mg kg–1 for 50 mg L–1 and 100 mg L–1, respectively). Manganese levels peaked with 100 mg L–1 ZnO NPs (28.6 ± 0.33 mg kg–1) and both ZnSO4 treatments (24.4 ± 0.38 and 28.6 ± 0.37 mg kg–1 for 50 mg L–1 and 100 mg L–1, respectively), demonstrating concentration--dependent mineral uptake dynamics.
Table 6. Zinc biofortification effects on mineral composition in flowers of dahlia cv. ‘Yaretzi’.
| Treatment (mg L–1) | Macronutrients (g kg–1) | ||||||
| N-total | P | K+ | Ca2+ | Mg2+ | |||
| Control | 20.3 ± 0.58a | 2.7 ± 0.05c | 8.1 ± 0.26c | 1.8 ± 0.03d | 2.2 ± 0.05c | ||
| 50ZnO NPs | 21.3 ± 0.78a | 2.8 ± 0.02b | 11.3 ± 0.36b | 2.2 ± 0.05a | 2.6 ± 0.02b | ||
| 100ZnO NPs | 21.1 ± 0.81a | 3.1 ± 0.06a | 12.9 ± 1.30a | 2.2 ± 0.05a | 2.7 ± 0.06a | ||
| 50ZnSO4 | 20.4 ± 0.44a | 2.8 ± 0.04b | 10.9 ± 0.42b | 1.8 ± 0.02c | 2.6 ± 0.03b | ||
| 100ZnSO4 | 20.7 ± 0.30a | 2.8 ± 0.02b | 11.8 ± 0.51a,b | 2.0 ± 0.04b | 2.3 ± 0.03c | ||
| Micronutrients (mg kg–1) | |||||||
| Fe2+ | Cu2+ | Mn2+ | Zn2+ | Na+ | B | ||
| Control | 49.7 ± 1.80c | 9.2 ± 1.04b | 21.4 ± 0.33b | 36.2 ± 1.55c | 13.6 ± 0.04c | 17.2 ± 0.93c | |
| 50ZnO NPs | 63.2 ± 3.73a | 12.8 ± 0.61a | 20.4 ± 0.28b | 43.5 ± 1.68b | 15.9 ± 0.07a | 23.3 ± 2.20a | |
| 100ZnO NPs | 55.3 ± 3.20b | 9.8 ± 0.83b | 28.6 ± 0.33a | 49.7 ± 0.57a | 15.8 ± 0.1a | 21.9 ± 3.11a,b | |
| 50ZnSO4 | 49.9 ± 1.39c | 13.0 ± 0.25a | 24.4 ± 0.38a | 42.4 ± 1.88b | 14.3 ± 0.08b,c | 19.9 ± 0.63b | |
| 100ZnSO4 | 59.4 ± 1.41a,b | 12.1 ± 0.34a,b | 28.6 ± 0.37a | 41.3 ± 0.62b | 15.0 ± 0.1b | 20.1 ± 0.62b | |
Notes: Mean values ± standard deviation (n = 4); values with the same superscript alphabets within each column do not differ statistically (P ≤ 0.05; Tukey’s test). Data is expressed on a DW basis.
ZnO NPs: zinc oxide nanoparticles; ZnSO4: zinc sulphate.
Pearson’s correlation analysis
Pearson’s correlation analysis (Table 7) revealed significant relationships between mineral content and quality attributes in dahlia flowers. Strong negative correlations (P < 0.05) were observed between: total N and TFl, r = –0.396); P and TA (r = –0.441) or TFl (r = –0.794); K+ and TFl (r = –0.560); Ca2+ and TA (r = –0.603), total phenols (r = –0.357), or TFl (r = –0.334); Mg2+ and TA (r = –0.582) or TAn (r = –0.405); Zn2+ and TA (r = –0.311) or TAn (r = –0.770); Fe2+ and TP (r = –0.612); and Cu2+ with TAn (r = –0.305), TFl (r = –0.392), or AA, r = –0.334). Conversely, significant positive correlations emerged between: total N and TA (r = 0.590); K+ and TP (r = 0.379); Mg2+ and TSS (r = 0.327) or AA (r = 0.372); Fe2+ and TSS (r = 0.314) or TFl (r = 0.480); Cu2+ and TP (r = 0.569); Zn2+ and TSS (r = 0.486) or AA (r = 0.378); and Na+ with TFl (r = 0.365) or AA (r = 0.453).
Table 7. Mineral–bioactive compound relationships in Zn-treated dahlia cv. ‘Yaretzi’ edible flowers.
| Mineral | TSS | TA | TAn | TP | TFl | AA |
|---|---|---|---|---|---|---|
| N-total | –0.233 | 0.590** | 0.262 | 0.259 | –0.396* | 0.120 |
| P | –0.296 | –0.441* | –0.133 | 0.238 | –0.794** | 0.093 |
| K+ | –0.058 | 0.154 | –0.125 | 0.379 | –0.560** | 0.234 |
| Ca2+ | 0.084 | –0.603** | –0.252 | –0.357* | –0.334* | –0.034 |
| Mg2+ | 0.327* | –0.582** | –0.405* | –0.172 | –0.582** | 0.372* |
| Fe2+ | 0.314 | –0.063 | 0.065 | –0.612** | 0.480* | 0.034 |
| Mn2+ | –0.322 | 0.133 | 0.045 | 0.001 | 0.097 | 0.254 |
| Cu2+ | 0.274 | 0.114 | –0.305* | 0.569** | –0.392* | –0.334* |
| Zn2+ | 0.486* | –0.311* | –0.770** | –0.121 | –0.270 | 0.378* |
| Na+ | 0.298 | –0.223 | –0.345* | 0.234 | 0.365* | 0.453* |
| B | 0.113 | –0.219 | 0.031 | 0.286 | 0.423* | 0.019 |
Notes: TSS: total soluble solids (°Brix); TA: titratable acidity (% citric acid); TAn: total anthocyanins (mg C3G 100 g–1); TP: total phenols (mg GAE g-1); TFl: total flavonoids (mg QE 100 g-1), AA: ascorbic acid (mg AA 100 g-1); GAE: gallic acid equivalent; QE: quercetin equivalent; C3G: cyanidin-3-glucoside.
*Significant at 5%, **significant at 1%. ns: not significant.
Discussion
Particularly in Mesoamerican cultures, EFs have held a prominent place in culinary traditions since ancient times. Here, species such as dahlia are prized for their cultural symbolisms, ornamental values, and culinary applications (Ghosh et al., 2025; Granados-Balbuena et al., 2023; Mejía-Muñoz et al., 2020). In recent years, the cultural and culinary heritages associated with dahlia’s EFs have gained renewed attention because of their (i) exceptional nutritional profile, being rich in bioactive phytochemicals (phenols, flavonoids, and anthocyanins), essential micronutrients and vitamins; (ii) low caloric density, making them ideal as components in health--conscious diets; and (iii) appealing sensory attributes (vibrant colours, diverse textures, and distinctive aromas) (Marchioni et al., 2024; Mulík et al., 2024). All these align with modern gastronomic trends seeking novel food experiences (Calderón-Jurado et al., 2023; Ojeda-Barrios et al., 2026; Rivera-Espejel et al., 2019). These characteristics make EFs particularly compatible with contemporary dietary patterns that emphasise plant-based nutrition and sustainable food production. Moreover, their high concentrations of antioxidant compounds and functional biomolecules enhance their nutritional value and are directly associated with health benefits, including metabolic regulation, prevention of chronic diseases, and body weight management (Abril-Carvajal et al., 2022; Reyes-Santiago et al., 2024).
This study provides the first scientific evidence that foliar applications of ZnO NPs significantly enhance chromaticity values (P ≤ 0.05) of dahlia flowers, establishing a novel biofortification approach for this and other EF species. Our findings yield three key contributions: (1) -biological—modulated pollinator attraction (bees/humming birds) through Zn-induced chromatic shifts, leveraging their well-documented spectral sensitivity (Chittka and Wells, 2004; Luizzi et al., 2021); (2) -agronomic—optimised nutrient delivery via ZnO NPs, offering superior efficiency over conventional methods; and (3) -ecological—potential enhancement of pollination services through improved floral visibility. The increased chromaticity directly correlates with hedonic value (Yoshida et al., 2009), a critical factor in premium EF markets, where colour drives ~70% of consumer preference. Comparative colour metrics contextualise our results: D. variabilis (L* = 50.17, C* = 34.59, and h° = 351.25; Costa et al., 2022) and peach flowers (e.g. ‘July Flame’: L* = 62.36 ± 1.78, C* = 13.49 ± 1.04, h° = 65.38 ± 0.63; Calderón-Jurado et al., 2023) demonstrate the competitive chromatic advantage of biofortified dahlias.
Photosynthetic pigments and leaf area serve as fundamental indicators of growth and development in plants, including dahlia (Martínez-Damián et al., 2025). Chlorophyll levels drive photosynthetic efficiency and biomass production, while carotenoids provide photoprotection against oxidative stress (Kusumiyati et al., 2025; Mallick et al., 2024; Villegas-Olguín et al., 2023). Together, they underpin plant vigour and are central to the visual appeal of many dahlia species cultivated for ornamental purposes (Bădulescu et al., 2023). Foliar application of Zn (100 mg L–1 ZnO NPs, and 50 mg L–1 and 100 mg L–1 ZnSO4) significantly enhanced Chl b and TC content in dahlia cv. ‘Yaretzi’ flowers, indicating improved Zn bioavailability in reproductive tissues (pistils, stamens, and petals). This micronutrient acts as a crucial cofactor for photosynthetic enzymes, including carbonic anhydrase and superoxide dismutase (Cakmak, 2000; Cakmak et al., 2017) while simultaneously triggering carotenoid biosynthesis as a photoprotective response under prolonged exposure to light (Granados-Balbuena et al., 2023). The accumulated carotenoids (lutein, β-carotene, and lycopene) not only mitigate oxidative stress but also generate characteristic yellow-to-red pigmentation in Asteraceae EFs, including dahlias, marigolds, and chrysanthemums (Kusumiyati et al., 2025; Ohmiya, 2011). Comparative analyses reveal species-specific responses to Zn supplementation: while gladiolus ‘White Prosperity’ showed increased chlorophyll (SPAD 54–64) with ZnSO4 (Karimian and Samiei, 2023), Eustoma ‘Mariachi Blue’ achieved higher total chlorophyll (13.27 µg mg–1 FW) with ZnO NPs (Seydmohammadi et al., 2019). Notably, dahlia carotenoid levels (8.01–8.90 µg β-carotene g–1 FW) were substantially lower than those in Tagetes spp. (2,843–3,998 µg 100 g–1 DW), highlighting important interspecific differences in these culturally significant Mesoamerican flowers (Mulik and Ozuna, 2020).
Petal colour of EFs is largely determined by the accumulation of fat- and water-soluble pigments, particularly carotenoids, flavonoids, and anthocyanins (Xu et al., 2020). In the present study, foliar application of 100 mg L–1 ZnO NPs significantly enhanced total anthocyanin content in dahlia flowers, suggesting a response associated with pigmentation and antioxidant metabolism in floral tissues. This response may be particularly relevant in heliophilous species, such as dahlia, whose phenotypic plasticity enables them to thrive under intense light conditions (Granados-Balbuena et al., 2024a; Villegas-Olguín et al., 2023). Anthocyanins are the main flavonoid-derived pigments responsible for red, purple, and blue hues in petals, including those of dahlia cultivars (Granados-Balbuena et al., 2024b). Recent studies on the dahlia cultivars ‘La Baron’ (dark pink to purple) and ‘Colorado Classic’ (light pink to bright lilac) have quantified their anthocyanin contents as 1.250 ± 0.002% DW and 1.138 ± 0.002% DW, respectively, with derivatives identified as delphinidin, cyanidin, petunidin, peonidin, and malvidin (Gontova et al., 2024). In addition, our results were consistent with previous reports for cultivated dahlia flowers, where anthocyanin concentrations ranged from 4.01 to 25 mg C3G 100 g–1 FW (Costa et al., 2022; Rivera-Espejel et al., 2019).
Our study demonstrates that Zn biofortification significantly increased TP, TFl, and AA contents in dahlia flowers, with the exception of the 50 mg L–1 ZnSO4 treatment, which showed no significant effect on TFl accumulation, compared to the control. This suggests a threshold effect in the regulation of this metabolic pathway. Beyond the overall increases in TP, TFl and AA, the pattern observed is consistent with a dose-dependent modulation of phenylpropanoid network, whereby Zn supply may enhance phenolic and flavonoid biosynthesis up to an optimum, but plateau or differentially affect specific branches once a threshold is reached (Mahdavi et al., 2022). Such non-linear responses are biologically plausible, given Zn’s tight coupling to redox homeostasis and enzyme regulation: moderate inputs can stimulate antioxidant-related metabolism, whereas higher levels may shift signalling and feedback control, thereby constraining further accumulation in particular subclasses (Ali et al., 2025). Anthocyanins, a key flavonoid class in pigmented petals, are especially responsive to micronutrient-driven changes in redox balance and precursor availability; accordingly, shifts in AA likely reflect coordinated antioxidant protection and pigmentation in floral tissues (Ohno, 2025).
As a key micronutrient, Zn plays a crucial regulatory role in the shikimic acid and phenylpropanoid pathways, directly influencing the expression of genes responsible for the biosynthesis of secondary metabolites (Cakmak et al., 2017; Palacio-Márquez et al., 2023). This mechanism explains the observed accumulation of phenolic compounds, which serve dual functions: (1) as part of the plant’s antioxidant defence system against oxidative stress in high metabolically active floral tissues (Martínez-Damián et al., 2021; Singh et al., 2021), and (2) as modulators of organoleptic properties (flavour and astringency) that influence both pollinator attraction and nutritional quality (Umar et al., 2021). The effectiveness of Zn biofortification in enhancing secondary metabolites has been well-documented in economically important crops, such as wheat, beans, and tomatoes (Bautista-Diaz et al., 2021; Du et al., 2019), with similar patterns observed in our study. Specifically, we found results comparable to those reported by Fortis-Hernández et al. (2023) for TP (40.52 mg GAE 100 g–1 FW) and TFl (4.39 mg QE 100 g–1 FW), and consistent with the findings in Stevia rebaudiana by Velázquez-Gamboa et al. (2024), where ZnO NPs (75–100 mg L–1) induced similar metabolic responses. Observed variations between studies can be attributed to several factors: (1) tissue-specific metabolic profiles (leaves as primary metabolic centres, versus flowers specialised in reproductive functions) (Xu et al., 2020); (2) experimental parameters (application frequency, dosage, and analytical methods); and (3) inherent differences between species and cultivars (Mahdavi et al., 2022). These findings underscore the importance of optimising Zn biofortification protocols according to target species and desired metabolic outcomes.
The findings of this study are particularly relevant because Zn biofortification research has focused mainly on conventional crops, such as cereals, fruit trees, vegetables, and medicinal plants (Preciado-Rangel et al., 2021; Umar et al., 2021). In contrast, ornamental species have remained largely unexplored, despite the long culinary use of dahlia. In this context, our results provide evidence that foliar Zn applications can modify the nutritional and phytochemical profile of edible dahlia flowers, consistent with reports of Zn-mediated responses in other plant systems (Azeem et al., 2025; Fortis-Hernández et al., 2024). These findings provide a useful basis for future studies aimed at evaluating Zn biofortification in dahlia and other edible flower species under different agronomic conditions.
Antioxidant capacity of EFs depends largely on bioactive compounds, such as polyphenols, carotenoids, and ascorbic acid (Marchioni et al., 2024). Given the structural diversity of these phytochemicals, accurate antioxidant capacity assessment requires multiple analytical methods (Monroy-García et al., 2025), prompting our use of ABTS, FRAP (125.02–152.61 μmol Fe2+/g), and DPPH (73–76% inhibition) assays. Comparative analysis revealed that our FRAP values were substantially lower than those reported for pomegranate flowers (609.66–670.22 mmol Trolox/g FW; Zhang et al., 2023), although similar to Zn-treated common beans (75% DPPH inhibition; Sida-Arreola et al., 2017). Notably, lettuce biofortified with ZnSO4 (75 μM) achieved higher antioxidant capacity (114 μM Trolox/100 g FW; Preciado-Rangel et al., 2021) than our DPPH results, while ZnO NPs (20 mg L–1) enhanced antioxidant capacity of basil (0.05 μM Trolox/100 g FW; Fortis-Hernández et al., 2023). These interspecific variations highlight the need to explore wider ranges of Zn concentration (<50 mg L–1 and >100 mg L–1) to optimise trade-off between antioxidant induction and potential phytotoxicity (Rizvi et al., 2023). The methodological predominance of DPPH assays stems from the radical’s stability, commercial availability, and procedural simplicity (Karimian and Samiei, 2023). However, our multi-method approach provided a more comprehensive antioxidant capacity assessment, reflecting Zn’s dual role as an enzymatic cofactor (e.g., superoxide dismutase) and as a modulator of non-enzymatic antioxidant biosynthesis (Bala et al., 2019; Nsabimana and Jiang, 2011). This is particularly relevant, given a delicate balance between the metabolic benefits of Zn and its capacity to induce oxidative stress at supraoptimal concentrations.
The incorporation of unconventional EFs, such as dahlia into modern diets, offers promising nutritional benefits for health-conscious consumers seeking low-calorie, nutrient-dense foods (Costa et al., 2022). However, their high moisture content (≥88%) poses significant postharvest challenges, including rapid dehydration, microbial proliferation (fungal and bacterial pathogens), and consequently limited shelf-life (Monroy-García et al., 2025). Our study revealed that ZnO NP treatments further elevated moisture levels beyond the reference value for dahlia cv. ‘Yaretzi’ (88.73%; Martínez-Damián et al., 2025), potentially through Zn’s regulation of aquaporin activity and subsequent effects on cellular turgor pressure. This physiological mechanism facilitates water and solute (e.g. sucrose, minerals) accumulation, ultimately influencing tissue expansion during floral development (López-Cervantes et al., 2018; Saeed et al., 2013). Comparative analyses showed species-specific moisture variations: rose (Rosa × grandiflora) exhibited lower values (84.56 ± 0.12%), while marigold (Calendula -officinalis) maintained levels (89.34 ± 0.1%) comparable to dahlia (De Lima Franzen et al., 2019). These differences reflect inherent physiological adaptations and highlight the need for tailored postharvest technologies to preserve the structural integrity and bioactive content of EFs.
Our study demonstrated that foliar application of ZnO NPs (50–100 mg L–1) significantly improved key nutritional parameters in dahlia flowers, including protein, fibre, ash, carbohydrate content and caloric value—making them particularly valuable for gourmet cuisine and as functional foods. Comparative analysis revealed that conventional dahlia cultivation (without Zn biofortification) typically yields lower nutritional values: D. pinnata shows reduced ash (7.19 ± 0.08 g 100 g–1), carbohydrates (47.03 ± 4.21 g 100 g–1), and energy content (19.02 ± 3.07 kcal 100 g–1) (Costa et al., 2022), while maintaining higher crude protein (13.80 ± 0.54 g 100 g–1) and comparable fibre levels (28.97 ± 3.58 g 100 g–1). Similarly, dahlia × hortorum cultivars exhibited elevated carbohydrate percentages (64.3–67.0%) across various colour morphotypes (Rivera-Espejel et al., 2019). Notably, our Zn-biofortified flowers surpassed the lipid content reported for wild and cultivated dahlia species (0.2–1.6%; Lara-Cortés et al., 2014). These enhancements underscore Zn’s crucial physiological roles as: (1) a structural component of carbonic anhydrase (regulating CO2 fixation; Mahdavi et al., 2022), (2) a chlorophyll biosynthesis cofactor, and (3) a mediator of carbohydrate translocation (Escudero-Almanza et al., 2025). The observed nutritional variability also reflected genetic diversity within the Dahlia genus, resulting from both intentional breeding and natural hybridisation under a diversity of edaphoclimatic conditions (Carrasco-Ortiz et al., 2019; Fujimoto and Onozaki, 2025; Gatt et al., 1998; Şekerci and Gülşen, 2016). This genetic plasticity necessitates genotype-specific optimisation of biofortification protocols to maximise nutritional outcomes while maintaining ornamental and culinary quality.
Edible flowers represent a promising low-calorie source of essential minerals (P, K+, Ca2+, Mg2+, Mn2+, and Na+) to complement human nutrition (Araújo et al., 2019; Calderón-Jurado et al., 2023). This study demonstrated that agronomic biofortification with ZnO NPs under open-field conditions enhanced Zn accumulation in dahlia flowers (36.2–49.7 mg kg–1 DW), with ZnO NP treatments outperforming conventional ZnSO4 treatments. The superior efficacy of ZnO NPs stems from their nanoscale properties, viz. increased foliar contact surface and controlled Zn2+ release (Cakmak et al., 2017), enabling more efficient penetration without risk of phytotoxicity (Preciado-Rangel et al., 2021). These findings have particular relevance to ornamental edible species such as dahlia, where precise Zn dosing must preserve petal quality (Mejía-Muñoz et al., 2020). Comparative analyses revealed that dahlia’s mineral accumulation followed the patterns observed in other crops. Thus, amaranth shoots accumulated 98–115 mg kg–1 DW of Zn under similar treatments (Reshma and Meenal, 2022), while basil showed 19.9% increases (192 μg g–1 DW) with ZnSO4 (Ciriello et al., 2022). Notably, dahlia’s K+, Mg2+, Ca2+, and Na+ levels remained below those reported for marigold (Tagetes erecta), calendula, lavender, and pansy (Araujo et al., 2019), but levels exceeded those of iron, copper, and Zn in nasturtium and coriander. In this study, low doses of ZnO NP increased the Fe2+ concentration. The observed Fe2+ response was consistent with a dose-dependent competition for shared divalent cation transporters (e.g. IRT1) between Zn and Fe. At a low dose (50 mg L–1), ZnO NPs may induce a non-specific upregulation of ZIP/YSL transporters, thereby enhancing concurrent Fe2+ uptake. However, higher doses (100 mg L–1) probably saturate these transport systems, favouring Zn uptake and consequently reducing Fe transport because of direct competition (Escudero-Almanza et al., 2025). Crucially, despite this competitive interaction at high Zn levels, dahlia flowers maintain a high basal Fe2+ content. This underscores the flower’s intrinsic richness in iron and suggests the presence of robust homeostatic mechanisms. These potentially include efficient metal chelation, vacuolar compartmentalisation, and potential stabilisation by ferredoxins, which collectively preserve iron bioavailability (White and Broadley, 2009; Wu et al., 2024). These variations reflect both genetic factors (differential expressions of ZIP, HMA, and MTP transporters in non-hyperaccumulators) and organ--specific allocation patterns in reproductive tissues (Sabzeali et al., 2025).
Plant development depends on the maintenance of a precise equilibrium between the availability of mineral nutrient and their complex interactions as metabolic cofactors and inhibitors. This is particularly true for the biosynthesis of secondary metabolites that mediate environmental adaptations (Ghosh et al., 2025). In EFs, these secondary compounds evolved primarily as pollinator attractants (Wink, 2010) and exhibit variable mineral bioavailability across floral structures (pistils, stamens, and petals) and developmental stages (Pinedo-Espinoza et al., 2020). While vegetative organs (leaves and stems) typically serve as primary nutrient reservoirs, with minerals often remobilised post-anthesis (Chahdoura et al., 2015), deficiencies during flowering can lead to severe reproductive impairment, including floral abortion or morphological deformity that compromise seed set (Brown et al., 2002). In this context, the mineral profile of flowers holds particular significance for human nutrition, as micronutrients, such as Zn, Fe, and Cu, play crucial roles in our cellular processes ranging from neurotransmission to enzymatic catalysis (Hou, 2022). Notably, the consumption of Zn-biofortified dahlia flowers may help to address widespread dietary Zn deficiencies, given this element’s involvement in more than 300 enzyme reactions in humans (Ahmed et al., 2024). This dual function (supporting both plant reproduction and human health) positions mineral-enhanced EFs as strategic components in sustainable food systems. The ephemeral nature of EFs demands careful management of nutrient allocation to optimise both agronomic yield and nutritional quality.
The safety of EFs, a growing functional food niche, is challenged by an underdeveloped regulatory framework, particularly regarding maximum residue limits of pesticides and heavy metals (Ojeda-Barrios et al., 2026; Sánchez-Chávez et al., 2026). From a food-safety perspective, use of ZnO NPs in EFs should be approached with caution because these products are commonly consumed fresh, which increases the relevance of dietary exposure to residual contaminants or nanoparticulate fractions, while external hazards in EFs may include both microbial and chemical impurities (Matyjaszczyk and Śmiechowska, 2019). Moreover, the safety of edible flower consumption remains under discussion because information on maximum safe daily intake is still limited and official safety requirements or guidelines are scarce (Dos Santos and Reis, 2021; Purohit et al., 2021). Thus, although ZnO NPs may improve Zn delivery, further studies are needed to determine Zn residues and nanoparticulate fractions in edible tissues and to assess dietary exposure, long-term consumption safety, and the possible transfer of nanoparticles through the food chain, which are not sufficiently interpreted (Sharma et al., 2025). Nevertheless, when EFs are produced under controlled conditions and handled hygienically, they may be considered a safe and attractive complement to the diet, providing bioactive compounds (e.g. polyphenols) and micronutrients of nutritional interest (Nicolau and Gostin, 2016). Surveillance data indicates that the main safety concerns of EFs are associated with microbial pathogens (e.g. Salmonella) and chemical contaminants, including pesticide residues, environmental pollutants, and potentially elevated metal concentrations (Carpena et al., 2024). For biofortified flowers, however, a proactive risk assessment remains essential, together with the standardisation of species-specific biofortification strategies, validated analytical protocols, and robust evidence on stability, bioaccessibility, and -consumer-relevant exposure.
Conclusions
Foliar application of ZnO NPs improved the nutritional and bioactive profile of dahlia EFs (cv. ‘Yaretzi’) and was more effective than conventional ZnSO4 under controlled open-field conditions. Specifically, 100 mg L–1 ZnO NPs increased several key quality attributes, including chroma, anthocyanin content (0.21 mg C3G 100 g–1), Zn concentration (49.7 mg kg–1), antioxidant capacity (152.61 µmol Fe2+ g–1), and proximate composition (e.g. 5.6% protein; and 30.3% fibre), without compromising floral appearance. Taken together, these findings support ZnO NPs as a promising nano--fertiliser approach to enhance the functional food potential of dahlia EFs. From a practical perspective, the results indicate that growers may use foliar ZnO NP programmes to increase micronutrient density and quality traits in EFs, provided that application rates are optimised and aligned with good agricultural practices to minimise off-target deposition. For food technologists, EFs should be managed as high-care fresh produce: rigorous hygiene, cold-chain control, and routine monitoring of microbiological hazards and chemical residues remain central to safeguarding safety and maintaining shelf quality. For nutritionists, EFs are best regarded as a complementary source of bioactive compounds and micronutrients rather than a principal vehicle for dietary Zn, given the small portions typically consumed as garnishes; nonetheless, they may add value within diversified diets. This work was limited to a single cultivar and a controlled production setting; therefore, further validation across cultivars, flower species, and commercial environments is warranted. Future research should address scalability, the long-term environmental fate of ZnO NPs (including potential soil accumulation and effects on non-target organisms), consumer acceptance of nano-biofortification, and additional safety evidence (e.g. in vitro toxicity and bioaccessibility assessments) to support responsible deployment.
Data Availability Statement
The data that supported the findings of this study is available from the corresponding author upon reasonable request.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Author Contributions
Conceptualisation: María Teresa Martínez-Damián and Damaris Leopoldina Ojeda-Barrios; methodology: Oscar Cruz-Alvarez, María del Rosario García-Mateos, and Yolanda Leticia Fernández-Pavía; validation: Oscar Cruz-Alvarez, Oscar Sariñana-Aldaco, and María del Rosario García-Mateos; formal analysis: Oscar Cruz-Alvarez and Orlanda Tanahiri García-González; investigation: Orlanda Tanahiri García-González, Oscar Cruz-Alvarez, and María del Rosario García-Mateos; resources, María Teresa Martínez-Damián and Damaris Leopoldina Ojeda-Barrios; data curation, Yolanda Leticia Fernández-Pavía and María Noemí Frías-Moreno; -writing—original draft preparation: Orlanda Tanahiri García-González and Oscar Cruz-Alvarez; writing—review and editing: María Teresa Martínez-Damián, Damaris Leopoldina Ojeda-Barrios, and Orlanda Tanahiri García-González; visualisation: Oscar Sariñana-Aldaco; supervision: María Teresa Martínez-Damián and Damaris Leopoldina Ojeda-Barrios; project administration: María Teresa Martínez-Damián; and fund acquisition: María Teresa Martínez-Damián. All authors read and agreed to the published version of the manuscript.
Funding
This work was jointly supported by the Universidad Autónoma Chapingo and the Universidad Autónoma de Chihuahua through institutional strategic project 25010-EI.
Acknowledgments
The authors thanked the Universidad Autónoma de Chihuahua for institutional support, and the staff of the laboratories at the Universidad Autónoma de Chapingo for their support in the analysis of this research.
Conflict of Interest
The authors declared no conflict of interest.
REFERENCES
Abril-Carvajal, L.M., Barona-Nieto, F.G., Hidalgo-Nuñez, D.R., Rodríguez-Cruz, V.J., and Vinueza-López C.N. 2022. Adición de flores comestibles en la elaboración de un embutido escaldado tipo pastel Mexicano. Ciencia Latina 6(5): 4015–4032. 10.37811/cl_rcm.v6i5.3373
Aguiñiga-Sánchez, I., Cadena-Íñiguez, J., Santiago-Osorio, E., Gómez-García, G., Mendoza-Núñez, V.M., Rosado-Pérez, J., Ruiz-Ramos, M., Cisneros-Solano, V.M., Ledesma-Martínez, E., Delgado-Bordonave, A.J., and Soto-Hernández, R.M. 2017. Chemical analyses and in vitro and in vivo toxicity of fruit methanol extract of Sechium edule var. nigrum spinosum. Pharmaceutical Biology 55(1): 1638–1645. 10.1080/13880209.2017.1316746
Ahmed, N., Zhang, B., Chachar, Z., Li, J., Xiao, G., Wang, Q., Hayat, F., Deng, L., Narejo, M.-u.-N., Bozdar, B., and Tu, P. 2024. Micronutrients and their effects on horticultural crop quality, productivity and sustainability. Scientia Horticulturae 323: 112512. 10.1016/j.scienta.2023.112512
Ali, S., Ali, B., Sajid, I.A., Ahmad, S., Yousaf, M.A., Ulhassan, Z., Zhang, K., Ali, S., Zhou, W., and Mao, B. 2025. Synergistic effects of exogenous melatonin and zinc oxide nanoparticles in alleviating cobalt stress in Brassica napus: insights from stress-related markers and antioxidant machinery. Environmental Science Nano 12: 368–387. 10.1039/D4EN00821A
Almendros, P., González, D., Fernández, M.D., García-Gómez, C., and Obrador, A. 2022. Both Zn biofortification and nutrient distribution pattern in cherry tomato plants are influenced by the application of ZnO nanofertilizer. Heliyon 8(3): e09130. 10.1016/j.heliyon.2022.e09130
American Dahlia Society (ADS). 2025. List of species dahlias. [Cited Jul. 21 2025]. Available at: https://www.dahlia.org/docsinfo/species-dahlias/list-of-species-dahlias/
Araújo, S., Matos, C., Correia, E., and Antunes, M.C. 2019. Evaluation of phytochemicals content, antioxidant activity and mineral composition of selected edible flowers. Quality Assurance and Safety of Crops & Foods 11(5): 471–478. 10.3920/QAS2018.1497
Association of Official Analytical Chemists (AOAC). 1990. Official Methods of Analysis, 14th ed. AOAC, Arlington, VA.
Azeem, A., Khan, S., Zia ul Haq, M., Shafiq, S., Aslam, M.T., Munir, S., and Sikandar Zaman, M. 2025. Biofortification strategies for enhancing crop nutritional value: a review of methods, challenges, and future directions. Discover Plants 2(1): 185. 10.1007/s44372-025-00276-3
Bădulescu, L., Badea, M.L., Ciobanu, M.I., and Toma, F. 2023. Morphological and physiological particularities of five dahlia cultivars. Scientific Papers. Series B. Horticulture 67(1): 495–503.
Bala, R., Kalia, A., and Dhaliwal, S.S. 2019. Evaluation of efficacy of ZnO nanoparticles as remedial zinc nanofertilizer for rice. Journal of Soil Science and Plant Nutrition 19: 379–389. 10.1007/s42729-019-00040-z
Bautista-Diaz, J., Cruz-Alvarez, O., Hernández-Rodríguez, O.A., Sánchez-Chávez, E., Jacobo-Cuellar, J.L., Preciado-Rangel, P., Ávila-Quezada, G.D., and Ojeda-Barrios, D.L. 2021. Zinc sulphate or zinc nanoparticle applications to leaves of green beans. Folia Horticulturae 33(2): 365–375. 10.2478/fhort-2021-0028
Benzie, I.F.F., and Strain, J.J. 1996. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: the FRAP assay. Analytical Biochemistry 239(1): 70–76. 10.1006/abio.1996.0292
Brand-Williams, W., Cuvelier, M.E., and Berset, C. 1995. Use of a free radical method to evaluate antioxidant activity. Food Science and Technology (LWT) 28(1): 25–30. 10.1016/S0023-6438(95)80008-5
Brown, P.H., Bellaloui, N., Wimmer, M.A., Bassil, E.S., Ruiz, J., Hu, H., Pfeffer, H., Dannel, F., and Römheld, V. 2002. Boron in plant biology. Plant Biology 4(2): 205–223. 10.1055/s-2002-25740
Cakmak, I. 2000. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytologist 146: 185–205. 10.1046/j.1469-8137.2000.00630.x
Cakmak, I., and Kutman, U.B. 2017. Agronomic biofortification of cereals with zinc: a review. European Journal of Soil Science (EJSS) 68(1): 1–16. 10.1111/ejss.12437
Cakmak, I., McLaughlin, M.J., and White, P. 2017. Zinc for-better crop production and human health. Plant Soil 411(1): 1–4. 10.1007/s11104-016-3166-9
Calderón-Jurado, M., Cruz-Alvarez, O., Juárez-López, P., Hernández-Rodríguez, O.A., Orozco-Meléndez, L.R., Morales-Maldonado, E.R., Ojeda-Barrios, D.L., and Alia-Tejacal, I. 2023. Bioactive compounds, minerals and antioxidants of edible flowers of peach and apple. International Journal of Food Properties 26(1): 1855–1866. 10.1080/10942912.2023.2236325
Carpena, M., Prieto, M.A., and Trząskowska, M. 2024. Chemical and microbial risk assessment of wild edible plants and flowers. EFSA Journal 22(S1): e221111. 10.2903/j.efsa.2024.e221111
Carrasco-Ortiz, M., Munguía-Lino, G., Castro-Castro, A., Vargas-Amado, G., Harker, M., and Rodríguez, A. 2019. Riqueza, distribución geográfica y estado de conservación del género Dahlia (Asteraceae) en México. Acta Botanica Mexicana 126: e1354. 10.21829/abm126.2019.1354
Chahdoura, H., Morales, P., Barreira, J.C.M., Barros, L., Fernández-Ruiz, V., Ferreira, I.C.F.R., and Achour, L. 2015. Dietary fiber, mineral elements profile and macronutrients composition in different edible parts of Opuntia microdasys (Lehm.) Pfeiff and Opuntia macrorhiza (Engelm.). Food Science and Technology (LWT) 64(1): 446–451. 10.1016/j.lwt.2015.05.011
Chang, C., Yang, M., Wen, H., and Chern, J. 2002. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. Journal of Food and Drug Analysis 10(3): 178–182. 10.38212/2224-6614.2748
Chetia, I., Vijayakumar, A., and Badwaik, L.S. 2025. Edible flowers’ flavor, safety and their utilization as functional ingredients: a review. Journal of Food Science and Technology 62: 11–23. 10.1007/s13197-024-06071-4
Chittka, L., and Wells, H. 2004. Color vision in bees: mechanisms, ecology and evolution. In: Prete, F.R. (ed.) Complex Worlds from Simpler Nervous Systems. MIT Press, Cambridge, MA, Ch. 7, pp. 165–191. 10.7551/mitpress/1994.003.0014
Ciriello, M., Formisano, L., Kyriacou, M., Soteriou, G.A., Graziani, G., De Pascale, S., and Rouphael, Y. 2022. Zinc biofortification of hydroponically grown basil: stress physiological responses and impact on antioxidant secondary metabolites of genotypic variants. Frontiers in Plant Science 13: 1049004. 10.3389/fpls.2022.1049004
Costa, P.A., Souza, D.C., Ossani, P.C., Mendes, M.H.A., Silva, M.L.S., Carvalho, E.E.N., and Resende, L.V. 2022. Nutritional and functional compounds in dahlia flowers and roots. Brazilian Journal of Food Technology (BJFT) 25: e2022029. 10.1590/1981-6723.02922
Davarpanah, S., Tehranifar, A., Davarynejad, G., Aran, M., Abadía, J., and Khorassani, R. 2017. Effects of foliar nano-nitrogen and urea fertilizers on the physical and chemical properties of pomegranate (Punica granatum cv. Ardestani) fruits. HortScience 52(2): 288–294. 10.21273/HORTSCI11248-16
De Lima Franzen, F., Rodríguez de Oliveira, M.S., Lidório, H.F., Farias Menegaes, J., and Martins Fries, L.L. 2019. Chemical composition of rose, sunflower and calendula flower petals for human food use. Ciencia Tecnology Agropecuaria 20(1): 149–168. 10.21930/rcta.vol20_num1_art:1252
Dos Santos, I.C., and Reis, S.N. 2021. Edible flowers: traditional and current use. Ornamental Horticulture 27(4): 438–445. 10.1590/2447-536x.v27i4.2392
Du, W., Yang, J., Peng, Q., Liang, X., and Mao, H. 2019. Comparison study of zinc nanoparticles and zinc sulphate on wheat growth: from toxicity and zinc biofortification. Chemosphere 227: 109–116. 10.1016/j.chemosphere.2019.03.168
Escudero-Almanza, D.J., Ojeda-Barrios, D.L., Cruz-Alvarez, O., Hernández-Rodríguez, O.A., and Ortiz-Rivera, Y. 2025. ZIP proteins related to zinc metabolism in plants. In: Tripathi, D.K., Singh, V.P., Pandey, S., Sharma, S., and Chauhan, D.K. (eds.) Zinc in Plants. Academic Press, Cambridge, MA, Ch. 8, pp. 173–192. 10.1016/B978-0-323-91314-0.00015-6
Fortis-Hernández, M., De la Rosa-Reta, C.V., Galindo-Guzmán, M., Preciado-Rangel, P., Galindo-Guzmán, A.P., and Flores-Loyola, E. 2023. Biofortification with ZnO NPs as nanofertilizers to improve sustainable commercial and phytochemical quality in basil plants. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 51(2): 13078. 10.15835/nbha51213078
Fortis-Hernández, M., Sánchez-Estrada, A., Hernández-Cruz, D., Lagunes-Fortiz, E., Betancourt-Galindo, R., and Fortiz-Hernández, J. 2024. Biofortification with zinc oxide nanoparticles via foliar increases biomass accumulation and lettuce phytochemical quality. Informacion Tecnica Economica Agraria (ITEA) 120(4): 344–359. 10.12706/itea.2024.009
Fujimoto, T., and Onozaki, T. 2025. Crossing and selection of dahlia (Dahlia Cav.) lines with a lower degree of petal abscission. Horticulture Journal 94(1): 15–23. 10.2503/hortj.QH-149
Gaines, T.P., and Mitchell, G.A. 1979. Boron determination in plant tissue by the azomethine H method. Communications in Soil Science and Plant Analysis 10(8): 1099–1108. 10.1080/00103627909366965
Gatt, M., Ding, H., Hammett, K., and Murray, B. 1998. Polyploidy and evolution in wild and cultivated Dahlia species. Annals of Botany 81(5): 647–656. 10.1006/anbo.1998.0614
Ghosh, P., Devraj, L., and Yadav, R.S. 2025. Edible flowers: potential applications for nutritional security. In: S.R. Purohit, V. Sharma, M., Kumari, K. Muthukumarappan and Kane-Potaka, J. Future Crops and Processing Technologies for Sustainability and Nutritional Security. (Eds.), CRC Press, Boca Raton, FL, Ch. 12, pp. 243–255.
Giusti, M.M., and Wrolstad, R.E. 2001. Characterization and -measurement of anthocyanins by UV-visible spectroscopy. In: R.E. Wrolstad (Ed.) Current Protocols in Food Analytical Chemistry. Wiley, New York, NY, Ch. F1.2, pp. 1-13. 10.1002/0471142913.faf0102s00
Gontova, T., Mashtaler, V., Romanova, S., Maloshtan, L., and Shanaida, M. 2024. Phytochemical analysis of anthocyanins extracted from the flowers of two dahlia cultivars and their-cytotoxic properties. Biomedical and Pharmacology Journal 17(2): 735–749. 10.13005/bpj/2900
Granados-Balbuena, S.Y., Chicatto-Gasperín, V., Santacruz-Juárez, E., Ocaranza-Sánchez, E., Ramírez-López, C., García-Meza, M.G., Tapia-López, L., Robles de la Torre, R.R., and Robles López, M.R. 2024a. Patented technologies in the extraction, preservation, and application of anthocyanins in food: a review. Applied Food Research 4(1): 100388. 10.1016/j.afres.2024.100388
Granados-Balbuena, S.Y., Díaz-Pacheco, A., García-Meza, M.G., Tapia-López, L., Cruz-Narváez, Y., and Ocaranza-Sánchez, E. 2023. Phytochemical profile of petals from black Dahlia pinnata by flow injection analysis–electrospray-ionization–Fourier transform ion cyclotron resonance mass spectrometry. Phytochemical Analysis 34(8): 1009–1021. 10.1002/pca.3268
Granados-Balbuena, S.Y., Gómez-Montaño, F.J., Pacheco, A.D., Ocaranza-Sánchez, E., and Canseco-González, D. 2024b. A concise review of Dahlia spp., modern and powerful techniques for anthocyanins detection. Chemistry Africa 7(7): 3549–3563. 10.1007/s42250-024-00972-9
Hatamian, M., Rezaie-Nejad, A., Kafi, M., Souri, M.K., and Shahbazi, K. 2018. Interactions of lead and nitrate on growth characteristics of ornamental judas tree (Cercis siliquastrum). Open Agriculture 3: 670–677. 10.1515/opag-2018-0070
Hernández-Epigmenio, F., García-Mateos, M.R., Sosa-Montes, E., Mejía-Muñoz, J.M., Fernández-Pavía, Y.L., Cruz-Alvarez, O., and Martínez-Damián, M.T. 2022. Phenolic profile and nutritional value of Dahlia × hortorum flowers. Revista Chapingo Serie Horticultura 28(3): 161–174.
Hou, T. 2022. Bioactive compounds in mineral bioavailability: activities, structures, and mechanisms. Frontiers in Nutrition 9: 1050670. 10.3389/fnut.2022.1050670
Hou, J., Wu, Y., Li, X., Wei, B., Li, S., and Wang, X. 2018. Toxic effects of different types of zinc oxide nanoparticles on algae, plants, invertebrates, vertebrates and microorganisms. Chemosphere 193: 852–860. 10.1016/j.chemosphere.2017.11.077
Irigoyen, J.J., Einerich, D.W., and Sánchez-Díaz, M. 1992. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiologia Plantarum 84(1): 55–60. 10.1111/j.1399-3054.1992.tb08764.x
Kah, M., Johnston, L.J., Kookana, R.S., Bruce, W., Haase, A., Ritz, V., Dinglasan, J., Doak, S., Garelick, H., and Gubala, V. 2021. Comprehensive framework for human health risk assessment of nanopesticides. Nature Nanotechnology 16: 955–964. 10.1038/s41565-021-00964-7
Karimian, Z., and Samiei, L. 2023. ZnO nanoparticles efficiently enhance drought tolerance in Dracocephalum kotschyi through altering physiological, biochemical and elemental contents. Frontiers in Plant Science 14: 1063618. 10.3389/fpls.2023.1063618
Khadanga, V., and Mishra, P.C. 2024. A review on toxicity mechanism and risk factors of nanoparticles in respiratory tract. Toxicology 504: 153781. 10.1016/j.tox.2024.153781
Kumari, P., Raju, D.V.S., Prasad, K.V., Singh, K.P., Saha, S., Arora, A., and Hossain, F. 2017. Quantification and correlation of-anthocyanin pigments and their antioxidant activities in rose (Rosa hybrida) varieties. Indian Journal of Agricultural Science 87(10): 1340–1346. 10.56093/ijas.v87i10.74991
Kusumiyati, Putri I.E., Hadiwijaya, Y., Kartika, A., Maulana, Y.E., and Sutari, W. 2025. Quality assurance of total carotenoids and-quercetin in marigold flowers (Tagetes erecta) as edible flowers. International Journal of Food Science 2025(1): 3277288. 10.1155/ijfo/3277288
Lara-Cortés, E., Martín-Belloso, O., Osorio-Díaz, P., Barrera-Necha, L.L., Sánchez-López, J.A., and Bautista-Baños, S. 2014. Antioxidant capacity, nutritional and functional composition of edible dahlia flowers. Revista Chapingo Serie Horticultura 20(1): 101–116. 10.5154/r.rchsh.2013.07.024
Liu, X., Xu, Y., Wang, Y., Yang, Q., and Li, Q. 2022. Rerouting-artificial light for efficient crops production: a review of lighting strategy in PFALs. Agronomy 12(5): 1021. 10.3390/agronomy12051021
López-Cervantes, J., Sánchez-Machado, D.I., Cruz-Flores, P., Mariscal-Domínguez, M.F., Mora-López, G.S., and Campas-Baypoli, O.N. 2018. Antioxidant capacity, proximate composition, and lipid constituents of Aloe vera flowers. Journal of Applied Research on Medicinal and Aromatic Plants 10: 93–98. 10.1016/j.jarmap.2018.02.004
Luizzi, V.J., Friberg, M., and Petrén, H. 2021. Phenotypic plasticity in floral scent in response to nutrient, but not water, availability in the perennial plant Arabis alpina. Functional Ecology 35(8): 1655–1665. 10.1111/1365-2435.13866
Mahdavi, S., Karimi, R., and Valipouri-Goudarzi, A. 2022. Effect of nano zinc oxide, nano zinc chelate and zinc sulfate on vineyard soil Zn-availability and grapevines (Vitis vinifera L.) yield and quality. Journal of Plant Nutrition 45(13): 1961–1976. 10.1080/01904167.2022.2046081
Mallick, S.R., Hassan, J., Hoque, M.A., Sultana, H., Kayesh, E., Ahmed, M., Ozaki, Y., Al-Hashimi, A., and Siddiqui, M.H. 2024. Color, proximate composition, bioactive compounds and antinutrient profiling of rose. Scientific Reports 14(1): 21690. 10.1038/s41598-024-72424-w
Marchioni, I., Rodolfi, M., Najar, B., Ruffoni, B., Machado, J., and Pistelli, L. 2024. Phytonutritional compounds and antioxidant activity of eight new edible flowers. Natural Product Research 38(21): 3728–3734. 10.1080/14786419.2023.2261140
Marcos-Gómez, R., Vera-Guzmán, A.M., Pérez-Ochoa, M.L., Martínez-Martínez, L., Hernández-Delgado, S., Martínez-Sánchez, D., and Chávez-Servía, J.L. 2025. Mineral content of four Mexican edible flowers growing in natural conditions and backyards from indigenous communities. Applied Sciences 15(7): 3432. 10.3390/app15073432
Martínez-Damián, M.T., López-Santiz, J.A., Hernández-Epigmenio, F., Sosa-Montes, E., and Cruz-Alvarez, O. 2025. Morphological and nutritional description of Yaretzi: a variety of dahlia for cut or potted flower. Revista Mexicana de Ciencias Agricolas16(4): e3689. 10.29312/remexca.v16i4.3689
Martínez-Damián, M.T., Mejía-Muñoz, J.M., Colinas-León, M.T., Hernández-Epigmenio, F., and Cruz-Alvarez, O. 2021. Nutritional value, bioactive compounds and capacity antioxidant in edible flowers of dahlia. Acta Scientiarum Polonorum Hortorum Cultus 20(5): 63–72. 10.24326/asphc.2021.5.6
Matyjaszczyk, E., and Śmiechowska, M. 2019. Edible flowers. Benefits and risks pertaining to their consumption. Trends Food Science and Technology 91: 670–674. 10.1016/j.tifs.2019.07.017
Mejía-Muñoz, J.M., De Luna-García, I., Jiménez-Ruiz, E.F., Sosa-Montes, E., Flores-Espinosa, C., Treviño-De Castro, G., and Reyes-Santiago, J. 2020. Research on dahlia, the national flower of Mexico. Acta Horticulturae 1288: 103–108. 10.17660/ActaHortic.2020.1288.15
Mejía-Muñoz, J., Flores-Espinosa, C., Peña-Ortega, G., Alvarado-Cano, M.E., Gaspar-Hernández, R., and Reyes-Santiago, J. 2015. Interspecific hybridization between Dahlia dissecta and D. rupicola. Acta Horticulturae 1087: 321–324. 10.17660/ActaHortic.2015.1087.41
Mera-Ovando, L.M., and Bye-Boettler, R. 2006. La Dahlia: una belleza originaria de México. Revista Digital Universitaria 7(11): 1–11.
Minnocci, A., Francini, A., Romeo, S., Sgrignuoli, A.D., Povero, G., and Sebastiani, L. 2018. Zn-localization and anatomical changes in leaf tissues of green beans (Phaseolus vulgaris) following foliar application of Zn-lignosulfonate and Zn EDTA. Scientia Horticulturae 231: 15–21. 10.1016/j.scienta.2017.12.002
Monroy-García, I.N., González-Galván, L.L., and Viveros-Valdez, E. 2025. Edible flowers as a source of bioactive -compounds. In: Segura Campos, M.R. (Ed.) Improving Health and Nutrition through Bioactive Compounds: Benefits and Applications. Elsevier, Amsterdam, the Netherlands, Ch. 11, p. 169–190. 10.1016/B978-0-443-21873-6.00012-9
Mulík, S., Hernández-Carrión, M., Pacheco-Pantoja, S.E., and Ozuna, C. 2024. Endemic edible flowers in the Mexican diet: understanding people’s knowledge, consumption, and experience. Future Foods 9: 100374. 10.1016/j.fufo.2024.100374
Mulík, S., and Ozuna, C. 2020. Mexican edible flowers: cultural background, traditional culinary uses, and potential health benefits. International Journal of Gastronomy and Food Science 21: 100235. 10.1016/j.ijgfs.2020.100235
Najar, B., Pieracci, Y., Fratini, F., Pistelli, L., Turchi, B., Varriale, D., Pistelli, L., Bozzini, M.F., and Marchioni, I. 2024. Exploring the volatile composition and antibacterial activity of edible flower hydrosols with insights into their spontaneous emissions and essential oil chemistry. Plants 13(8): 1145. 10.3390/plants13081145
Nicolau, A.I., and Gostin, A.I. 2016. Safety of edible flowers. In: Prakash, V., Martín-Belloso, O., Keener, L., Astley, S.B., Braun, S., McMahon, H., and Lelieveld, H. (Eds.) Regulating Safety of Traditional and Ethnic Foods. Academic Press, Cambridge, MA, pp. 395–419. 10.1016/B978-0-12-800605-4.00021-9
Niu, J., Liu, C., Huang, M., Liu, K., and Yan, D. 2021. Effects of foliar fertilization: a review of current status and future perspectives. Journal of Soil Science and Plant Nutrition 21: 104–118. 10.1007/s42729-020-00346-3
Nsabimana, C., and Jiang, B. 2011. The chemical composition of some garden Dahlia tubers. British Food Journal 113(9): 1081–1093. 10.1108/00070701111174541
Ohmiya, A. 2011. Diversity of carotenoid composition in flower-petals. Japan Agricultural Research Quarterly (JARQ) 45(2): 163–171. 10.6090/jarq.45.163
Ohno, S. 2025. Flavonoid biosynthesis in Dahlia (Dahlia variabilis) flowers. Horticulture Journal 94(4): 429–437. 10.2503/hortj.SZD-R007
Ojeda-Barrios, D.L., Martínez-Damián, M.T., Hernández-Rodríguez, O.A., Martínez-Ruiz, A., and Cruz-Alvarez, O. 2026. Phytochemicals and biological activities of flowers of dahlia. In: Murthy, H.N. (Ed.) Bioactive Compounds in Edible Flowers. CRC Press, Boca Raton, FL, Ch. 7, pp. 163–188. 10.1201/9781003645979-9
Ontiveros-Capurata, R.E., Juárez-López, P., Mendoza-Tafolla, R.O., Alia-Tejacal, I., Villegas-Torres, O.G., Guillén-Sánchez, D., and Cartmill, A.D. 2022. Relationship between chlorophyll and nitrogen concentration, and fresh matter production in basil ‘Nufar’ (Ocimum basilicum) with three handheld chlorophyll meter readings: SPAD, at LEAF and MC-100. Revista Chapingo Serie Horticultura 28(3): 189–202. 10.5154/r.rchsh.2021.09.018
Palacio-Márquez, A., Ramírez-Estrada, C.A., Sánchez, E., and Chávez-Mendoza, C. 2023. Biofortification with zinc: the use of nanoparticles as an alternative to improve the nutritional quality of crops. In: Chen, J.T. (Ed.) Advanced Nanotechnology in Plants: Methods and Applications. CRC Press, Boca Raton, FL, Ch. 6, pp. 1–17.
Pinedo-Espinoza, J.M., Gutiérrez-Tlahque, J., Santiago-Saenz, Y.O., Aguirre-Mancilla, C.L., Reyes-Fuentes, M., and López-Palestina, C.U. 2020. Nutritional composition, bioactive compounds and antioxidant activity of wild edible flowers consumed in semiarid regions of Mexico. Plant Foods for Human Nutrition 75: 413–419. 10.1007/s11130-020-00822-2
Preciado-Rangel, P., Campos-Ortiz, A., Sánchez-Chávez, E., Reyes-González, A., Ruiz-Espinoza, F., Ojeda-Barrios, D., and Hernández-Montiel, L. 2021. Zinc biofortification improves yield, nutraceutical quality and antioxidant capacity in lettuce. Tropical and Subtropical Agroecosystems 24(3): 122. 10.56369/tsaes.3844
Purohit, S.R., Rana, S.S., Idrishi, R., Sharma, V., and Ghosh, P. 2021. A review on nutritional, bioactive, toxicological properties and preservation of edible flowers. Future Foods 4: 100078. 10.1016/j.fufo.2021.100078
Rameshraddy, G., Pavithra, G.J., Rajashekar Reddy, B.H., Salimath, M., Geetha, K.N., and Shankar, A.G. 2017. Zinc oxide nano particles increases Zn uptake, translocation in rice with positive effect on growth, yield and moisture stress tolerance. Indian Journal of Plant Physiology 22: 287–294. 10.1007/s40502-017-0303-2
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., and Rice-Evans, C. 1999. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biology and Medicine 26(9–10): 1231–1237. 10.1016/S0891-5849(98)00315-3
Reshma, Z., and Meenal, K. 2022. Foliar application of biosynthesised zinc nanoparticles as a strategy for ferti-fortification by improving yield, zinc content and zinc use efficiency in amaranth. Heliyon 8(10): e10912. 10.1016/j.heliyon.2022.e10912
Reyes-Santiago, J., Ortiz-Brunel, J.P., Lichter-Marck, I.H., and Castro-Castro, A. 2024. Another new species of the genus Dahlia (Asteraceae, Coreopsideae): are we close to knowing its total diversity? Acta Botanica Mexicana 131: e2270.
Rivera-Espejel, E.A., Cruz-Alvarez, O., Mejía-Muñoz, J.M., García-Mateos, M.R., Colinas-León, M.T., and Martínez-Damián, M.T. 2019. Physicochemical quality, antioxidant capacity and nutritional value of edible flowers of some wild dahlia species. Folia Horticulturae 31(2): 331–342. 10.2478/fhort-2019-0026
Rizvi, A., Saleem, S., Solanki, B., Ahmed, B., Singh, R., and Khan, M.S. 2023. Nano-biofortified crop plants with zinc for human health. In: Rajput, V.D., El-Ramady, H., Upadhyay, S.K., Minkina, T., Ahmed, B., and Mandzhieva, S. (Eds.) Nano-Biofortification for Human and Environmental Health. Springer, Cham, Switzerland, Ch. 4, pp. 53–76. 10.1007/978-3-031-35147-1_4
Sabzeali, F., Ahmadikhah, A., Farrokh, N., and Haghi, R. 2025. Genome-wide identification and transcription profiling of safflower (Carthamus tinctorius) HD-ZIP gene family under water deficit. BMC Genomics 26: 874. 10.1186/s12864-025-12060-4
Saeed, T., Hassan, I., Jilani, G., and Abbasi, N.A. 2013. Zinc augments the growth and floral attributes of gladiolus, and alleviates oxidative stress in cut flowers. Scientia Horticulturae 164: 124–129. 10.1016/j.scienta.2013.09.017
Sánchez-Chávez, E., Munguía-Lino, G., López-Pérez, J.D., and Morales-Ramírez, S. 2026. Beyond ornamentals: ethnobotany of Mexican Dahlia (Asteraceae), the national flower of Mexico. Botanical Science 104(2): 1–19. 10.17129/botsci.3827
Şekerci, A.D., and Gülşen, O. 2016. Overview of dahlia breeding. Scientific Papers. Series B. Horticulture 60: 199–204.
Seydmohammadi, Z., Roein, Z., and Rezvanipour, S. 2019. Accelerating the growth and flowering of Eustoma grandiflorum by foliar application of nano-ZnO and nano-CaCO3. Plant Physiology Reports 25(1): 140–148. 10.1007/s40502-019-00473-9
Sharma, V., Maurya, A., Kandhol, N., Singh, V.P., Sharma, S., Peralta-Videa, J., and Tripathi, D.K. 2025. Zinc deficiency in plants: an insight into fortification strategies. In: Tripathi, D.K., Singh, V.P., Pandey, S., Sharma, S., and Chauhan, D.K. (Eds.) Zinc in Plants. Academic Press, Cambridge, MA, Ch. 10, pp. 249–264. 10.1016/B978-0-323-91314-0.00011-9
Sida-Arreola, J.P., Sánchez, E., Ojeda-Barrios, D.L., Ávila-Quezada, G.D., Flores-Córdova, M.A., Márquez-Quiroz, C., and Preciado-Rangel, P. 2017. Can biofortification of zinc improve the antioxidant capacity and nutritional quality of beans? Emirates Journal of Food and Agriculture 29(3): 237–241. 10.1080/23311932.2017.1283725
Silveira, A.C., Vilaró, F., Kvapil, M.F., Rodríguez, S.D.C., and Zaccari, F. 2020. Physical–chemical characterization and potential for frying of genetic potato (Solanum tuberosum) materials. Revista Chapingo Serie Horticultura 26(2): 143–157. 10.5154/r.rchsh.2019.09.015
Singh, V., Singh, J., Kushwaha, R., Singh, M., Kumar, S., and Rai, A.K. 2021. Assessment of antioxidant activity, minerals and chemical constituents of edible mahua (Madhuca longifolia) flower and fruit using principal component analysis. Nutrition & Food Science 51(2): 387–411. 10.1108/NFS-04-2020-0129
Sokal, R.R., and Rohlf, F.J. 1995. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd edition. W.H. Freeman, New York, NY.
Tomasik, P. 2003. Saccharides and polysaccharides: an introduction. In: Tomasik, P. (Ed.) Chemical and Functional Properties of Food Saccharides. CRC Press, Boca Raton, FL, Ch. 1, p. 1–19. 10.1201/9780203495728
Umar, W., Hameed, M.K., Aziz, T., Maqsood, M.A., Bilal, H.M., and Rasheed, N. 2021. Synthesis, characterization and application of ZnO nanoparticles for improved growth and Zn biofortification in maize. Archives of Agronomy and Soil Science 67(9): 1164–1176. 10.1080/03650340.2020.1782893
Velázquez-Gamboa, M.C., Rodríguez-Hernández, L., Abud-Archila, M., Gutiérrez-Miceli, F.A., Ruíz-Valdiviezo, V.M., Valdez-Salas, B., and Luján-Hidalgo, M.C. 2024. Effect of zinc and magnesium oxide phytonanoparticles on the production of rebaudiosides and phenolic compounds in Stevia rebaudiana Bertoni plants. Biotecnia 26: e2313. 10.18633/biotecnia.v26.2313
Villegas-Olguín, M.A., Mendoza-Villarreal, R., Benavides-Mendoza, A., García-Osuna, H.T., Cabrera-de la Fuente, M., and Robledo-Torres, V. 2023. Agronomic response of four Dahlia pinnata Cav. (Asteraceae) varieties in three production environments. Agrociencia 57(8): 1–13. 10.47163/agrociencia.v57i8.2946
Wang, H., Xu, D., Jiang, F., Wang, S., Wang, A., Liu, H., Lei, L., Qian, W., and Fan, W. 2024. The genomes of Dahlia pinnata, Cosmos bipinnatus, and Bidens alba in tribe Coreopsideae provide insights into polyploid evolution and inulin biosynthesis. GigaScience 13: giae032. 10.1093/gigascience/giae032
Wellburn, A.R. 1994. The spectral determination of chlorophyll A and chlorophyll B as well as total carotenoids using various solvents with spectrophotometers of different resolution. Journal of Plant Physiology 144(3): 307–313. 10.1016/S0176-1617(11)81192-2
White, P.J., and Broadley, M.R. 2009. Biofortification of crops with seven mineral elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, selenium, and iodine. New Phytologist 182(1): 49–84. 10.1111/j.1469-8137.2008.02738.x
White, P.J., and Broadley, M.R. 2011. Physiological limits to zinc biofortification of edible crops. Frontiers in Plant Science 2: 80. 10.3389/fpls.2011.00080
Whitley, G.R. 1985. The medicinal and nutritional properties of Dahlia spp. Journal of Ethnopharmacology 14(1): 75–82. 10.1016/0378-8741(85)90031-5
Wink, M. 2010. Introduction: biochemistry, physiology and -ecological functions of secondary metabolites. In: Wink, M. (Ed.) Annual Plant Reviews, Vol. 40: Biochemistry of Plant Secondary Metabolism. Blackwell, Oxford, UK, Ch. 1, pp. 1–19. 10.1002/9781444320503.ch1
Wu, M., Bian, X., Huang, B., Du, Y., Hu, S., Wang, Y., Shen, J., and Wu, S. 2024. HD-Zip proteins modify floral structures for self-pollination in tomato. Science 384(6691): 124–130. 10.1126/science.adl1982
Xu, K., Zhang, M., Fang, Z., and Wang, B. 2020. Degradation and regulation of edible flower pigments under thermal processing: a review. Critical Reviews in Food Science and Nutrition 61(6): 1038–1048. 10.1080/10408398.2020.1752142
Yoshida, K., Mori, M., and Kondo, T. 2009. Blue flower color-development by anthocyanins: from chemical structure to cell physiology. Natural Product Reports 26(7): 884–915. 10.1039/b800165k
Zhang, H., Wang, M., Yu, G., Pu, J., Tian, K., Tang, X., Du, Y., Wu, H., Hu, J., Luo, X., Lin, L., and Deng, Q. 2023. Comparative analysis of the phenolic contents and antioxidant activities of different parts of two pomegranate (Punica granatum L.)-cultivars: ‘Tunisia’ and ‘Qingpi’. Frontiers in Plant Science 14: 1265018. 10.3389/fpls.2023.1265018
Zhu, J., Li, J., Shen, Y., Liu, S., Zeng, N., Zhan, X., White, J.C., and Gardea-Torresdey, J. 2020. Mechanism of zinc oxide nanoparticle entry into wheat seedling leaves. Environmental Science Nano 7(12): 3901–3913. 10.1039/D0EN00658K