
Download

Total anthocyanin content m g / L = A × M W × D F × 1000 ε × I
RESEARCH ARTICLE
Phytochemical composition, antioxidant potential, and bulb mass of differently colored onions under distinct sowing methods
Đorđe Vojnović1, Žarko Ilin1,2, Aleksandar Ž. Kostić3, Boris Adamović1, Sofija Kilibarda3, Lato Pezo4, Jelena Popović-Đorđević3*
1Faculty of Agriculture, University of Novi Sad, Dositej Obradović Square 8, Novi Sad, Serbia;
2Academy of Engineering Sciences of Serbia, Kraljice Marije 16, Belgrade, Serbia;
3Faculty of Agriculture, University of Belgrade, Belgrade, Serbia;
4Institute of General and Physical Chemistry, University of Belgrade, Studentski Trg 12–16, Belgrade, Serbia
Abstract
Onion (Allium cepa L.), either fresh or processed, is widely consumed for its unique aroma, flavor, and taste. While farmers aim to maximize yield, consumers demand high-quality products. These goals can conflict under certain cultivation conditions but can be balanced through appropriate agronomic practices. Therefore, this study aimed to investigate the effects of two sowing methods, the standard sowing method (SSM) and the new sowing method (NSM), characterized by reduced seed rates and wider row spacing on selected quality parameters (onion bulb weight and total soluble sugars [TSS]), key bioactive compounds content, total phenolic content (TPC), total flavonoid content (TFC), hydroxycinnamic acids (HCAs), total monomeric anthocyanins (TMA), and antioxidant properties of 10 new onion genotypes (G1–G10). The results revealed statistically significant differences (p < 0.005) among both sowing methods and genotypes, indicating that these factors substantially impact the measured parameters. Bright violet, skin-colored genotype G8 stood out with the highest values of TPC, TFC, HCAs, TMA, TSS, and antioxidant activity (measured in three assays) in the NSM and bulb weight in the SSM. These findings suggest that the NSM may offer economic benefits for farmers, while selected genotypes provide consumers with high-quality onions.
Key words: Allium cepa L, genotype, quality, soluble sugars, secondary metabolites, flavonoids, monomeric anthocyanins, antioxidant activity
*Corresponding Author: Jelena Popović-Đorđević, Faculty of Agriculture, University of Belgrade, 11080 Belgrade, Serbia. Email: jelenadj@agrif.bg.ac.rs
Academic Editor: Theodoros Varzakas, PhD, Department Food Science and Technology, Food Technology and Food Safety Management Systems, University of Peloponnese, Antikalamos, 24100 Kalamata, Greece
Received: 22 February 2025; Accepted: 23 April 2025; Published: 1 July 2025
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Onion (Allium cepa L.), a member of the Alliaceae family, is one of the most essential vegetable species worldwide. Allium species are cultivated and consumed worldwide, with onion being the most economically significant species in the Balkan region and Serbia. Vuković et al. (2022) highlighted its leading role in vegetable production and market share in this region. Onion is widely used in culinary applications (bulbs, leaves, or whole plants), and it is consumed raw, fried, roasted, or pickled to enhance the flavor of various dishes. Consumed worldwide, onion is valued for its aroma, and flavor, as well as for richness of bioactive compounds such as vitamins, total phenols, flavonoids, and anthocyanins (Bibi et al., 2022; Kim et al., 2024; Ren and Zhou, 2021; Vuković et al., 2023a). Different onion colors are closely linked to varying levels of bioactive compounds, influencing both consumer preferences and market value. Red and violet onions, for example, are particularly rich in flavonoids, especially quercetin and anthocyanins, which contribute to their intense color and strong health benefits (Kim et al., 2024; Ren and Zhou, 2021). Such characteristics increase their appeal for export markets and premium product lines.
In order to cure and treat some disorders but to avoid synthetic drugs, plants have been recognized as an excellent source of natural medicines both for human (Chaachouay and Zidane, 2021; Ivanz et al., 2018) as well as animal welfare (Kuralkara and Kuralkar, 2021). Since ancient times, onions have been used in traditional medicine to treat inflammation and common cold (Sagar et al., 2022; Vuković et al., 2023a). These medicinal properties are attributed to a wide array of bioactive compounds, including organosulfur compounds and phenolic compounds (Vuković et al., 2023a; Zhao et al., 2021). The aroma, pungency, and numerous health effects of this species are mostly associated with these compounds, particularly sulfur-containing substances (such as cysteine sulfoxides and thiosulfinates), as well as flavonoids (quercetin and its glycosides), anthocyanins, and phenolic acids (Metrani et al., 2020; Sagar et al., 2022; Teshika et al., 2019). These bioactive constituents are responsible for the antioxidant, antibacterial, antiviral, and anticancer activities of onions, supporting their extensive use in traditional and modern medicine.
The goal of onion producers (farmers) is to achieve the highest yield of first-class onion bulbs, as they command the highest market price. Meanwhile, consumers prioritize a higher content of bioactive compounds because of their numerous health benefits. Therefore, it is essential to advance onion production to achieve both high yields and nutritional quality. In pursuit of these goals, numerous studies have focused on optimizing irrigation methods (Geries et al., 2021; Piri and Naserin, 2020), fertilization practices (Golubkina et al., 2022; Rafie et al., 2017), biostimulant applications (Vojnović et al., 2024; Vuković et al., 2023b), and pesticide management (Degani and Kalman, 2021; Nault et al., 2012). Such agronomic factors directly influence the phytochemical composition of onion bulbs by affecting plant stress levels, nutrient availability, and metabolic activity, which are key drivers in the synthesis of phenolic compounds, flavonoids, and anthocyanins. However, studies on the impact of direct sowing methods on onion yield and bulb quality remain limited (Ketema et al., 2018; Jeevitha et al. 2024). This is primarily because open-field direct sowing involves a lengthy germination period—lasting up to 30 days (Vojnović et al., 2024)—during which weeds and drought may threaten crop establishment. Despite these challenges, direct sowing remains highly attractive to farmers, as it reduces production costs and labor inputs compared to transplanting, while also allowing for higher plant density and, consequently, the potential for increased yields. Therefore, further studies on direct sowing under practical, open-field conditions are essential to provide farmers with reliable guidelines for achieving stable yields and high bulb quality, while optimizing economic efficiency in onion production. Among onions grown for fresh consumption, bronze-skinned genotypes are the most common. Recent studies, however, emphasize the importance of food color as a critical sensory cue that influences taste expectations (Spence, 2015). The genetic diversity of onions provides an excellent opportunity to enhance color variety in foods, with a wide range of bulb colors available (white, gold, red, yellow, bronze, etc.) (Chalbi et al., 2023; Schwinn et al., 2016). Although numerous studies have assessed onion genotype diversity in controlled, pot-based experiments (Alam et al., 2023; Arena et al., 2024; Corrado et al., 2024; Gökçe et al., 2023), only few have investigated the performance of these genotypes under practical, open-field conditions, which is necessary for results that are applicable to real-world farming.
The genotype of a vegetable can significantly influence the content of its bioactive compound. For instance, Chalbi et al. (2023) found that red-skinned onion genotypes contain higher levels of total phenols than white-skinned ones. Onion is an excellent example of how genotype can impact both flavonoid content and antioxidant activity (Chalbi et al., 2023), with red genotypes typically containing more flavonoids than the yellow variety (Ren and Zhou, 2021). Gorrepati et al. (2024) recently reported that anthocyanins were the most abundant phenolic compounds that highly influenced the color of the onion peel in short-day dark red genotypes. In addition, Chalbi et al. (2023) noted that the onion genotype significantly affects sugar synthesis, an essential factor in determining bulb flavor.
Given the diverse colors, content of bioactive compounds, and nutritional value of the onions, there is a need to improve current onion bulb production practices to achieve high yields and good quality for both domestic and export markets. Although some studies have examined the effects of planting methods (Alemu et al., 2022; Alkhateeb et al., 2024; Gebretsadik and Dechassa, 2018) and onion genotypes (Alam et al., 2023; Arena et al., 2024; Bibi et al., 2022; Gökçe et al., 2023) in onions grown from seedlings, there is no enough research on the effects of genotype in relation to onions grown from direct sowing. This gap is particularly significant, as farmers predominantly grow onions through direct sowing.
In response to this scientific gap, the objective of this study was to examine the impact of direct sowing methods on the selected quality parameters (bulb weight and soluble sugars content), content of bioactive compounds, and antioxidant properties in new onion genotypes with different skin colors.
Materials and Methods
Study site
This study investigated ten onion genotypes under two sowing methods, in a field experiment that was conducted in 2022 in the village of Gospođinci, an important vegetable-growing region in the Republic of Serbia. The environmental conditions at the study site, characterized by a moderate continental climate, are representative of major onion-producing regions in Europe and other temperate zones worldwide (Arena et al., 2024; Golubkina et al., 2022), making the findings relevant for broader agricultural practice. The field (45.24 N, 19.59 E) used for this experiment has been cultivated by local farmers for vegetable production over an extended period. Before establishing the experiment, soil samples were collected at a depth of 30 cm using a probe (50 mm diameter) and analyzed in a laboratory for soil, fertilizer, and plant material testing at the Faculty of Agriculture Novi Sad. Based on the analysis, the soil pH was 7.17 (which was appropriate for onion cultivation), and the CaCO3 content was 5.7%. The humus content was 1.8%, with a nitrogen (N-min) content of 51.8 kg/ha. The phosphorus pentoxide (P2O5) content was slightly below the optimal level (12.76 mg/100 g soil), while the potassium oxide (K2O) content was high (43.2 mg/100 g soil), likely because of long-term intensive vegetable production on this soil. The field is also located near the Jergička River, whose water was used for irrigation during onion cultivation.
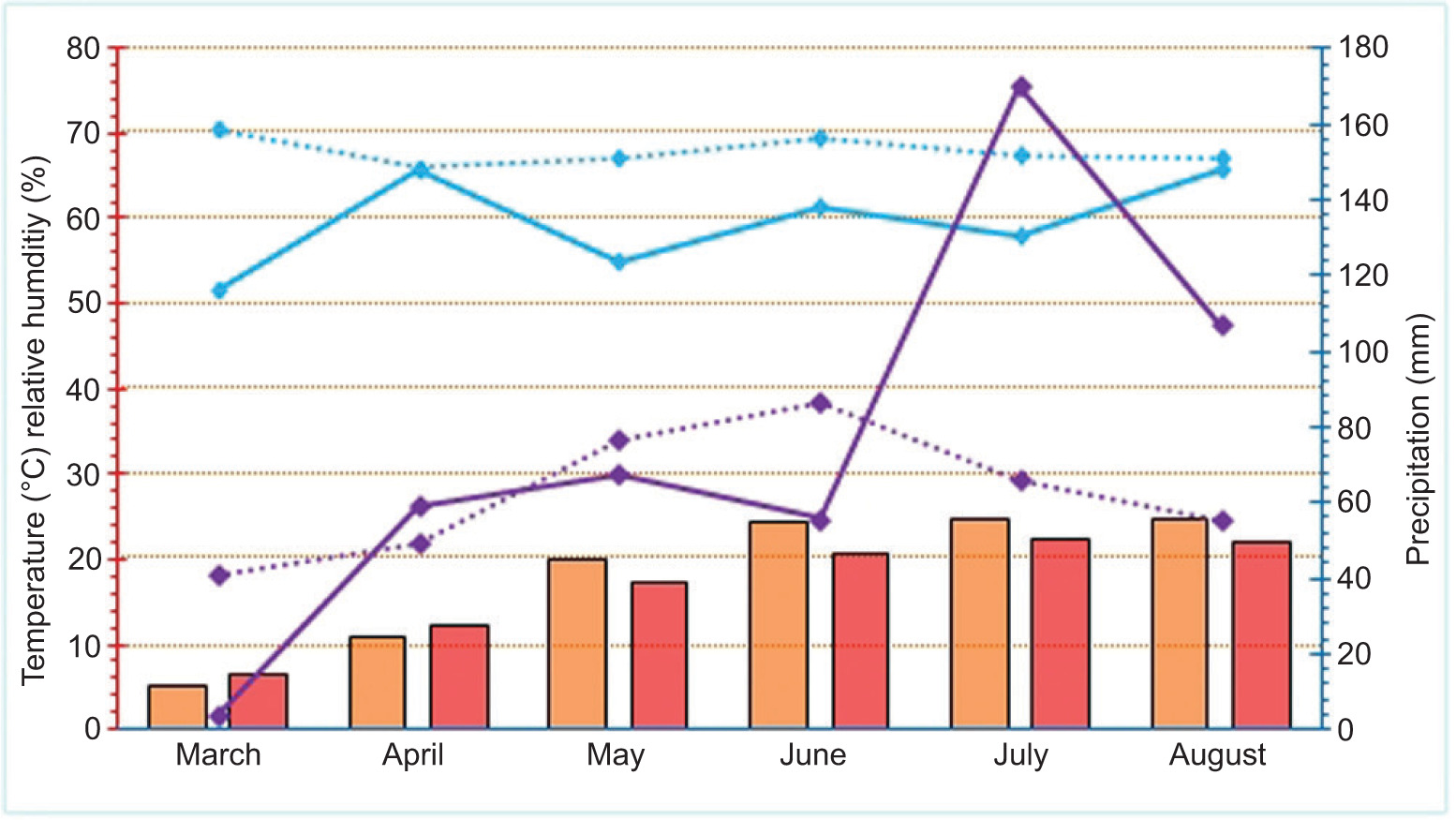
During the experiment, meteorological parameters were closely monitored with a digital weather station Vantage Pro2TM by Davis Instruments® (Hayward, CA, USA), which was powered by solar panels. In 2022, the lowest mean daily temperature was recorded in March (5.03°C), which was 1.58°C below the long-term average (1991–2020) (Figure 1). The highest daily temperature was recorded in July (24.8°C), exceeding the long-term average by 2.39°C. The lowest rainfall occurred in March (3.6 mm), which was 37.2 mm below the long-term average, while the highest rainfall was in July (169.8 mm), 103.9 mm above the long-term average (1991–2020). Heavy rainfall in July and August delayed bulb harvesting by 2–3 weeks. The lowest humidity was recorded in March (51.7%), while the highest was in April, showing a difference of 19.01% and 0.17% below the long-term average (1991–2020), respectively.
Figure 1. Meteorological parameters during the field experiment in 2022. The orange bars represent the mean daily temperatures, while the red bars indicate the multiyear average (1991–2020). The purple line represents the total precipitation, and the dashed purple line indicates the multiyear average of precipitation (1991–2020). The solid blue line represents the relative humidity, while the dashed blue line indicates the multiyear average of relative humidity (1991–2020).
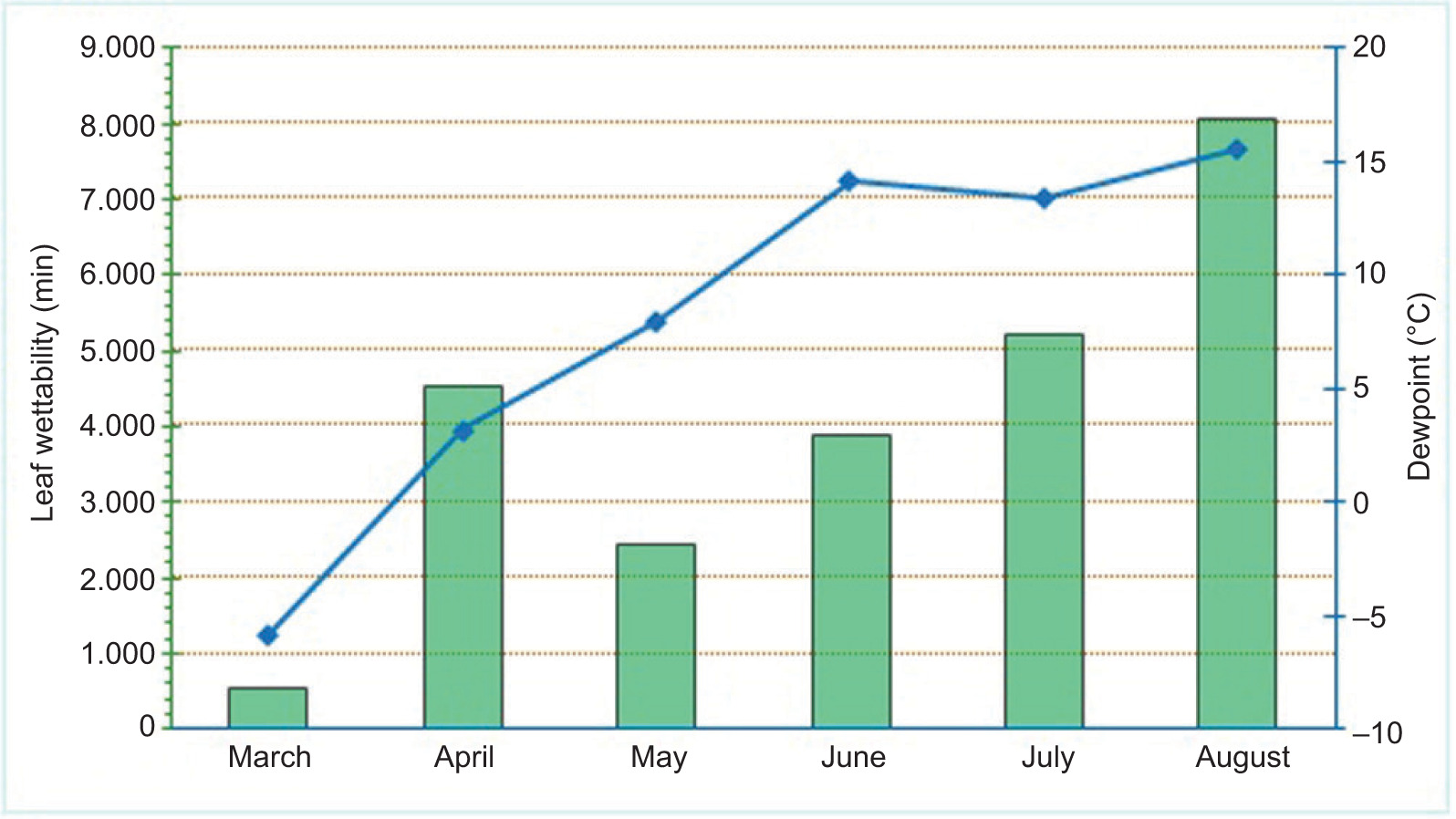
To improve the forecasting of plant disease, farmers increasingly rely on parameters such as dew point and duration of leaf wetness. During this study, the lowest average dew point was recorded in March (–5.80°C), and the highest was recorded in August (15.5°C) (Figure 2). The shortest duration of leaf wetness was in March (555.0 min), while the longest duration was in August (8060.0 min).
Figure 2. Duration of leaf wetness (green bars) and dew point (blue line) during the field experiment in 2022.
Experimental treatment
The experiment was organized following a split-plot method in a randomized block design with three replications. A split-plot design was chosen because randomizing sowing methods across small plots would have been logistically challenging and could have introduced errors in the precision of sowing. This design ensured optimal field organization and allowed for reliable statistical evaluation of the collected data. The main plots consist of two direct sowing methods for onion: (1) Standard sowing method (SSM)—planting in eight-row beds with four double rows per bed, with a spacing of 50 cm between beds, 20 cm between double rows, 10 cm between rows within each double row, and 5.53 cm between plants within a row and (2) New sowing method (NSM)—planting in six-row beds with three double rows per bed, also with 50 cm between beds, 35 cm between double rows, 10 cm between rows within each double row, and 5.53 cm between plants within a row.
The subplots consisted of 10 commercial onion genotypes (G1–G10), presented in Figure 3 and Table 1, selected with the cooperation of local producers—“Family farm Đukić” and “Family farm Salonski” from Gospođinci—who are among the largest onion growers in the region. These genotypes were chosen because they have not yet been evaluated under local open-field conditions. Farmers expressed interest in testing their performance, given the growing market demand for differently colored onion bulbs, particularly in urban areas where consumer preferences are shifting toward more diverse and visually appealing products The size of the main plot was 100 m2, and the size of the subplot was 10 m2 (1.5 m width × 6.67 m length), with the experiment conducted in three replicates. The total size of the field experiment was 600 m2 (two main plots × three replicates).
Figure 3. Studied onion genotypes: (A) Elenka F1, (B) Ambrador F1, (C) Magika F1, (D) Khalifer F1, (E) KNS 314 F1, (F) KNS 315 F1, (G) Fiamma F1, (H) Lisa F1, (I) Zelda F1, and (J) White Opera F1.
Table 1. Characteristics of onion genotypes.
| Genotype code | Genotype name | Primary characteristics | |||
|---|---|---|---|---|---|
| Skin color | Length of vegetation | Storability | Usage | ||
| G1 | 1Elenka F1 | Dark bronze | Medium late | Long storage | Fresh market |
| G2 | 1Ambrador F1 | Dark bronze | Medium late | Long storage | Fresh market/processing |
| G3 | 1Magika F1 | Bronze | Medium early | Long storage | Fresh market |
| G4 | 1Khalifer F1 | Bronze | Medium | Long storage | Fresh market |
| G5 | 2KNS 314 F1 | Light bronze | Early | Unknown | Fresh market/processing |
| G6 | 2KNS 315 F1 | Orange-bronze | Early | Unknown | Fresh market/processing |
| G7 | 1Fiamma F1 | Dark red/violet | Medium early | Long storage | Fresh market/processing/bunching |
| G8 | 1Lisa F1 | Bright violet | Medium | Long storage | Fresh market/processing |
| G9 | 1Zelda F1 | Light red | Medium | Long storage | Fresh market/processing |
| G10 | 1White Opera F1 | Bright white | Medium | Long storage | Fresh market/processing (freezing) |
Genotypes labeled as 1 are from Cora Seeds® (Cesena, Italy), while those labeled as 2 are from an unknown seed producer.
Agronomic practices
The preceding crop on the experimental plot was wheat (Triticum aestivum L.). After the wheat harvest on June 20th, the field was disked to a depth of 10 cm to break capillary water flow and retain soil moisture. In the third week of August, the field was treated with Glifosav 480 (Chemical Agrosava®, Šimanovci, Serbia) herbicide (glyphosate 480 g/L) at a rate of 5 L/ha. In the fall, 500 kg/ha of a compound NPK 15:15:15 (Yara®, Oslo, Norway) fertilizer was applied, providing 75 kg each of N, P, and K per hectare. On the same day, plowing was done to a depth of 30 cm using a Titan 140 s-cx plow by Regent® (Attnang-Puchheim, Austria). In early spring, harrowing was done to close the winter furrow and reduce evaporation from the soil surface. The first pre-sowing preparation was done with a combined cultivator 2 weeks before sowing. A week before sowing, N fertilizer urea (46% N) was applied at 200 kg/ha, providing 92 kg N/ha, followed by a second pre-sowing preparation. Direct sowing was performed using a pneumatic seed drill SNT-2/3-290 by Agricola Italiana® (Massanzago, Italy), equipped with seed plates containing 72 holes arranged in a single row, with a hole diameter of 1.1 mm. The sowing rate was controlled using a 22-tooth sprocket (A) and a 16-tooth sprocket (B), with the gear set at the 10° position. Simultaneously, rolling was conducted to improve seed-to-soil contact, using a roller attached to the tractor’s front hydraulic system. The direct sowing and rolling were completed on March 14th, and onion emergence was recorded 28 days later.
Based on measurements of the duration of leaf wetness and dew point, and in consultation with the experts from Agricultural Extension Service Sombor (2024, https://www.psssombor.rs/pis/), regular weed, disease, and pest control measures were implemented, as shown in Table S1. In addition, fertilizer and pesticide were applied according to the results of soil analysis and meteorological data, following precise expert recommendations, to ensure optimal crop development and to prevent the accumulation of harmful residues in onion bulbs.
The onion was irrigated through a drip irrigation system using 16 mm drip lines by Scarabeli Irrigazione® (Bologna, Italy), with 10 cm emitter spacing and a flow rate of 10 L/h/m. Solar panels were used as the energy source to power the irrigation pump. The timing of irrigation and the amount of water were determined using SKU-6440 sensors by Davis Instruments® (Hayward, CA, USA), connected to a weather station (Vojnović et al., 2024).
Measurement of the weight of onion bulbs
Samples were collected from each genotype, with 15 randomly selected plants per replication (45 per genotype). The same person collected samples from all treatments to reduce random error and obtain objective differences between treatments. Onion bulbs were brought to the vegetable laboratory at the Faculty of Agriculture in Novi Sad. The average bulb weight was calculated by measuring the total bulb mass from each plot and dividing it by the total number of bulbs, as per standard practice in onion bulb weight assessment. Later, bulbs were peeled, ground, and stored at -20°C until chemical analysis (Gorenje FH45ECW freezer, Velenje, Slovenia).
Extraction procedure
For chemical analyses, onion bulbs of different genotypes were defrosted and subjected to extraction process immediately after being transported to the chemical laboratory.
Extraction procedure for phenolic compounds: Bulbs were ground and homogenized in order to obtain a representative sample(s) per genotype/sowing method with a mass-solvent ratio of 1:10. In order to prevent protein extraction, 80% methanol was used as a solvent. In this way, only phenolic compounds were extracted preventing interferences during quantitative determinations. Aqueous methanol has been recognized as one of the most suitable extraction solvents because of adequate polarity that prevents degradation of phenolics and leads to the highest yield of extracted compound(s), as it was demonstrated for several vegetable agro wastes (Babbar et al., 2014). Extraction was performed for 3 hours with intensive shaking of cuvettes that were protected from the light during the extraction process. After the extraction, samples were centrifuged, and supernatants were separated from solid residue and stored in a refrigerator until further analysis was performed.
Extraction procedure for anthocyanins: Crushed and ground bulbs of three violet-colored genotypes (G7, G8, and G9) were extracted with 80% methanol acidified (0.1%) with concentrated HCl under the same conditions as it was for samples used for other analyses.
Phytochemical characterization of onion bulbs
In order to analyze selected phytochemicals and nutrients, spectrophotometric assays were used for the measurement of total phenolic content (TPC), total flavonoid content (TFC), total hydroxycinnamic acid (HCA) derivative content, total monomeric anthocyanins (TMA), and total soluble sugars (TSS).
Total phenolic content
In order to determine the TPC, the previously described method (Kostić et al., 2021) was employed with some modifications. Adequately diluted samples (180 µL) were mixed with working solution of Folin-Ciocalteu reagent (900 µL). After 5 min, 900 µL of sodium carbonate solution (7.5%) was added and the reaction mixture was left to stand for 90 min, protected from the light. Absorbance of finally developed color was read at 765 nm and was used for quantification. Results are expressed as mg/g gallic acid equivalents (GAE) based on the fresh weight (FW) of samples.
Total flavonoid content
In order to determine the TFC, 1 mL of the original (undiluted) extract was mixed with 1 mL of 2% AlCl3 (prepared in acidified methanol) in an Eppendorf cuvette. After keeping the samples in the dark for 10 min, the absorbance was read at 430 nm, as proposed in the literature (Chalbi et al., 2023).
Total hydroxycinnamic acid derivative content
In order to determine the content of HCAs, the previously described methodology was applied (Kostić et al., 2021). It implied mixing of 200 µL of properly diluted extracts with the same volumes (400 µL) of 0.5 M HCl, Arnow’s reagent, and 2.125 M NaOH solution, respectively. Furthermore, working solutions were diluted with 0.6 mL of deionized water (miliQ water) and left to react for 20 min. After developing of color, the absorbance of each sample was read at 525 nm in order to HCAs. Results are expressed as mg/g chlorogenic acid equivalents (CGAE) based on the FW of samples.
Total monomeric anthocyanin content
Description of method: for the determination of the total monomeric anthocyanin content, two buffer systems were prepared: 0.025 M potassium chloride (pH=1) and 0.4 M sodium acetate buffer (pH=4.5) as proposed in the literature (Rockenbach et al., 2012). In separate tubes, every sample was mixed with adequate buffer solution in a 1:1 ratio (v:v). Samples were mixed and left for 15 min for the development of color. The absorbance for both tubes was read at two different wavelengths: 520 nm and 700 nm. The resulting absorbance (A) for every sample was obtained according to the following calculation:
A= (A510 – A700) pH=1 – (A510 – A700) pH=4.5
The total content of anthocyanin was calculated according to the following equation:
where MW–a molar mass of cyanidin-3-glucoside, which is 449.2; DF–dilution factor; ε–molar absorptivity for cyanidin-3-glucoside, which is 26900; l–cuvette width, which is 1 cm.
The obtained results are expressed as mg/100 g of cyaniding-3-glucoside equivalents (C3GE) based on the FW of samples.
Total soluble sugar content
The content of TSS was determined according to the suggestions in the literature (Aluta et al., 2023; Dubois et al., 1956), with some modifications. Appropriately diluted extracts (200 µL) were mixed with the same volume of 5% phenol. After that, 3 mL of concentrated sulfuric acid was carefully added. The reaction mixture was left in the dark to react for 30 min. The developed color was measured at an absorbance of 490 nm. Results are expressed as mg/g of glucose based on the FW of samples.
Determination of the antioxidant activity of onion bulbs
The antioxidant properties of extracts obtained from onion bulbs were evaluated with the application of four standard antioxidant assays: in vitro phosphomolybdenum total antioxidant capacity (TAC), cupric reducing antioxidant capacity (CUPRAC), quenching of 2,2-diphenyl-1-picrylhydrazyl radical (DPPH•), and quenching of 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid radical cation (ABTS•+), as described in the literature (Kostić et al., 2023). Analyses were done in triplicates, and results were expressed as mg/g ascorbic acid equivalents (AAE) for TAC and CUPRAC methods, whereas for DPPH• and ABTS•+ assays, results were expressed as µmol/g Trolox equivalents (TE), all based on FW of the onion samples.
Statistical analysis
The data were statistically analyzed using the STATISTICA 10.0 software package (StatSoft Inc., Tulsa, OK, USA), with all measurements conducted in triplicate. Results are presented as mean values accompanied by their standard deviations (SD). Prior to further analysis, all samples were tested for variance homogeneity using Levene’s test and for normal distribution using the Shapiro–Wilk test, chosen for their robustness and suitability for agricultural datasets. To assess the differences in measured parameters among onion treatments, analysis of variance (ANOVA) was performed. Tukey’s HSD post hoc test was then applied to compare the means and determine statistically significant variations between the samples. In addition, color correlation analysis was conducted to examine the similarity in the content of active compound across different treatments, allowing for a more detailed comparison of their chemical compositions. The Pearson color correlation analysis was performed using R software version 4.0.3 (64-bit). The correlation between variables was visualized using a color-coded correlation matrix, where red tones indicate negative correlations, blue tones indicate positive correlations, and the intensity of the color reflects the strength of the correlation. The diameter of the circles is proportional to the absolute value of the correlation coefficient, providing a dual representation of correlation strength.
Cluster analysis (CA) and Principal Component Analysis (PCA) were employed to explore the chemical parameters derived from various onion treatments. The CA and PCA plots provide an insightful overview, revealing patterns and trends that enhance the understanding of the characteristic profiles of the observed samples.
Results
Phytochemical characteristics and the weight of onion bulbs
The results of bulb weight and chemical parameters (TPC, TFC, HCAs, TMA, and TSS), along with antioxidant potential (TAC, CUPRAC, DPPH•, and ABTS•+) for 10 genotypes (G1–G10) under two distinct sowing methods, SSM and NSM, are presented in Table 2. The F-test results indicate statistically significant differences among groups, both for sowing methods and genotype types. The p-values (p<0.001) suggested that these differences were significant at the 99.9% confidence level, confirming that both the sowing method and genotype had a significant impact on the measured parameters.
Table 2. Studied parameters for 10 onion genotypes.
| Sowing | Genotype | TPC mg/g GAE | TFCmg/g QE | HCAsmg/g GAE | TMAmg/100 g C3GE | TSSmg/g Glc | TACmg/g AAE | CUPRAC mg/g AAE | DPPHµmol/g TE | ABTS•+μmol/g TE | Bulb weightg |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SSM | G1 | 1.06±0.05e | 0.50±0.01g | n.d.** | n.d. | 84.81±20.10abcde | 4.88±0.32abcde | 0.55±0.02g | 2.23±0.15cdef | 9.65±0.07e | 135.79±13.05cdefg |
| SSM | G2 | 1.00±0.11de | 0.44±0.01fg | n.d. | n.d. | 111.93±0.14e | 6.55±0.42fg | 0.53±0.02fg | 2.20±0.13cdef | 4.24±0.20bcd | 139.03±3.90defg |
| SSM | G3 | 0.70±0.03bc | 0.34±0.00ef | n.d. | n.d. | 87.58±4.70abcde | 5.27±0.24abcdef | 0.15±0.01ab | 1.93±0.02abcdef | 3.92±0.14abcd | 118.91±13.26bcdefg |
| SSM | G4 | 0.71±0.04bcd | 0.27±0.00cde | n.d. | n.d. | 93.78±15.66bcde | 6.08±0.31cdefg | 0.27±0.04bcde | 1.81±0.20abcde | 4.34±0.01bcd | 148.27±18.19fg |
| SSM | G5 | 0.61±0.05bc | 0.18±0.00abcd | n.d. | n.d. | 75.43±1.63abcde | 6.67±0.61fg | 0.19±0.02abc | 2.06±0.15bcdef | 3.34±0.29abcd | 58.88±4.12a |
| SSM | G6 | 0.53±0.03b | 0.18±0.01abcd | n.d. | n.d. | 102.39±15.93de | 6.29±0.35defg | 0.17±0.02ab | 1.48±0.07a | 3.65±0.59abcd | 94.56±5.47abcd |
| SSM | G7 | 0.60±0.05bc | 0.13±0.01a | 0.43±0.01d | 2.73±0.23b | 58.24±2.48ab | 4.60±0.34ab | 0.17±0.01ab | 2.45±0.15fgh | 4.95±0.57cd | 142.54±24.20efg |
| SSM | G8 | 0.80±0.01bcde | 0.22±0.00abcd | 0.36±0.02c | 5.84±0.46c | 90.23±1.41bcde | 6.60±0.03fg | 0.43±0.01defg | 2.85±0.18gh | 5.52±0.44d | 153.10±22.05g |
| SSM | G9 | 0.84±0.02cde | 0.34±0.00ef | 0.26±0.00b | 0.86±0.00a | 88.54±10.98abcde | 3.94±0.19a | 0.35±0.01bcdefg | 2.30±0.01defg | 5.44±0.60d | 130.78±6.00cdefg |
| SSM | G10 | 0.11±0.00a | n.d. | n.d. | n.d. | 61.21±3.98abc | 4.39±0.38ab | <LOQ | 1.76±0.22abcd | 1.72±0.02a | 155.37±24.15g |
| NSM | G1 | 0.71±0.05bcd | 0.26±0.00cde | n.d. | n.d. | 77.23±12.69abcde | 4.29±0.44a | 0.31±0.01bcdef | 1.73±0.06abc | 3.95±0.43abcd | 103.38±6.28abcdef |
| NSM | G2 | 0.70±0.08bcd | 0.35±0.09ef | 0.24±0.00a | n.d. | 57.80±8.51ab | 4.74±0.47abc | 0.29±0.02bcdef | 1.91±0.11abcdef | 3.88±0.42abcd | 135.79±8.21cdefg |
| NSM | G3 | 0.67±0.07bc | 0.24±0.00bcd | n.d. | n.d. | 49.15±3.14a | 4.84±0.05abcd | 0.21±0.02bcd | 1.69±0.01abc | 3.71±0.31abcd | 94.59±9.39abcd |
| NSM | G4 | 0.83±0.09cde | 0.41±0.01fg | n.d. | n.d. | 68.89±3.26abcd | 4.00±0.31a | 0.48±0.06efg | 1.61±0.09ab | 4.65±0.81cd | 143.06±22.14efg |
| NSM | G5 | 0.60±0.05bc | 0.15±0.00ab | n.d. | n.d. | 73.43±7.01abcde | 6.94±0.36g | 0.21±0.01abcd | 1.96±0.14abcdef | 3.27±0.01abcd | 78.59±11.84ab |
| NSM | G6 | 0.68±0.09bc | 0.26±0.00cde | n.d. | n.d. | 88.40±9.83abcde | 5.41±0.71abcdef | 0.27±0.03bcde | 1.75±0.11abcd | 2.88±0.14abc | 91.07±11.00abc |
| NSM | G7 | 0.77±0.01bcde | 0.16±0.01abc | 0.38±0.01c | 2.58±0.21b | 60.08±9.70abc | 5.80±0.25bcdefg | 0.42±0.05cdefg | 2.87±0.11h | 5.23±0.41cd | 139.02±24.35defg |
| NSM | G8 | 2.06±0.23f | 0.66±0.04h | 0.80±0.01e | 6.92±0.43d | 99.54±10.01cde | 6.34±0.43efg | 2.00±0.24h | 6.52±0.27i | 20.82±2.12f | 119.55±15.04bcdefg |
| NSM | G9 | 0.79±0.04bcde | 0.28±0.04de | 0.25±0.00ab | n.d. | 56.88±5.31ab | 4.43±0.28ab | 0.31±0.02bcdef | 2.32±0.18efgh | 4.40±0.37bcd | 98.42±9.49abcde |
| NSM | G10 | 0.16±0.01a | <LOQ* | n.d. | n.d. | 89.36±14.66bcde | 5.25±0.07abcdef | <LOQ | 2.00±0.05abcdef | 2.07±0.05ab | 143.40±9.91efg |
| F | 53.13 | 80.20 | 2293.45 | 339.70 | 6.48 | 14.10 | 97.16 | 119.03 | 89.38 | 10.35 | |
| p | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
Values with the same letter in a column are not significantly different at p<0.05 (Tukey’s HSD test); * LOQ–below the limit of quantification; **n.d.–not determined.
TPC–Total Phenolic Content (mg/g gallic acid equivalents (GAE)); TFC–Total Flavonoid Content (mg/g quercetin equivalents (QE)); HCAs–Hydroxycinnamic Acid derivatives (mg/g chlorogenic acid equivalents (CGAE)); TMA–Total Monomeric Anthocyanin (mg/100 g of cyaniding-3-glucoside equivalents (C3GE)); TSS–Total Soluble Sugars (mg/g of glucose); TAC–Total Antioxidant Capacity (mg/g ascorbic acid equivalents (AAE)) ; CUPRAC–Cupric Reducing Antioxidant Capacity (mg/g ascorbic acid equivalents (AAE)); DPPH•–Free Radical Scavenging Activity (μmol/g Trolox equivalents (TE)); ABTS•+–Antioxidant Activity by the ABTS•+ method (µmol/g Trolox equivalents (TE)). All results are expressed on fresh weight (FW) of onion bulb samples.
The highest TPC was observed in genotype G8 × NSM (2.06±0.23 mg/g GAE), which is statistically significantly higher than in all other genotypes. In contrast, genotype G10, under both SSM and NSM, demonstrated the lowest TPC values. The analysis confirmed that both the sowing method and genotype statistically significantly influenced TPC values (p<0.05).
For TFC, treatment G8 × NSM exhibited the highest value (0.66±0.04 mg/g QE), while genotype G7 under SSM had the lowest TFC value of 0.13±0.01 mg/g QE. A clear pattern of significant differences was observed across genotypes, and between sowing methods, with NSM frequently showing higher TFC values.
HCAs were measured in only three samples, genotypes G7–G9 under both sowing methods, and in genotype G2 × NSM. The highest content of HCAs was observed in G8 × NSM (0.80±0.01 mg/g CGAE), statistically significantly different from most other genotypes, while the lowest value of HCAs was recorded for G2 × NSM (0.24±0.00 mg/g CGAE).
Similar pattern was noticed for the content of TMA in onion samples. The highest TMA content was found in genotype G8 × NSM (6.92±0.43 mg/100 g C3GE), and the lowest in genotype G9 × SSM (0.86±0.00 mg/100 g C3GE). TMA concentrations varied statistically significantly between genotypes (p<0.05), with genotype G8 (both sowing methods) displaying exceptionally high values (Table 2).
Regarding TSS, genotype G2 under SSM stood out with the highest value (111.93±0.14 mg/g Glc), statistically significantly differing from the other genotypes (p<0.05). TSS values varied substantially among genotypes, but overall genotypes sowed in SSM tend to result in higher TSS values compared to NSM (Table 2).
Antioxidant characteristics of onion bulbs
Antioxidant properties of onion genotypes, measured by selected assays, differed between samples sowed under different methods (p<0.05). The highest values were found for G5 × NSM (6.94±0.36 mg/g AAE, TAC assay) and G8 × NSM (2.00±0.24 mg/g AAE, 6.52±0.27 µmol/g TE, and 20.82±2.12 µmol/g TE, CUPRAC, DPPH•, and ABTS•+ assays, respectively) (Table 2). The genotype G10 × SSM was among genotypes that expressed the lowest (TAC and ABTS•+) or no (CUPRAC) antioxidant potential in applied assays. TAC, CUPRAC, and DPPH• values of studied onion samples tend to be higher under NSM compared to SSM. The high DPPH• radical scavenging activity of the genotype G8 (6.52±0.27 µmol/g TE) indicated a strong antioxidant capacity for this genotype under NSM conditions.
Finally, bulb weight showed considerable variation, with genotype G10 under SSM having the highest weight (155.37±24.15 g), while genotype G5 under SSM recorded the lowest weight (58.88±4.12 g). However, G5 × SSM did not statistically significantly differ from genotypes G6 (SSM) and G1, G3, G5, G6, and G9 (NMS) with respect to this yield component parameter (p> 0.05). Significant variability in bulb weight was observed among genotypes and sowing methods, with both methods showing substantial differences (p<0.05) (Figure 4).
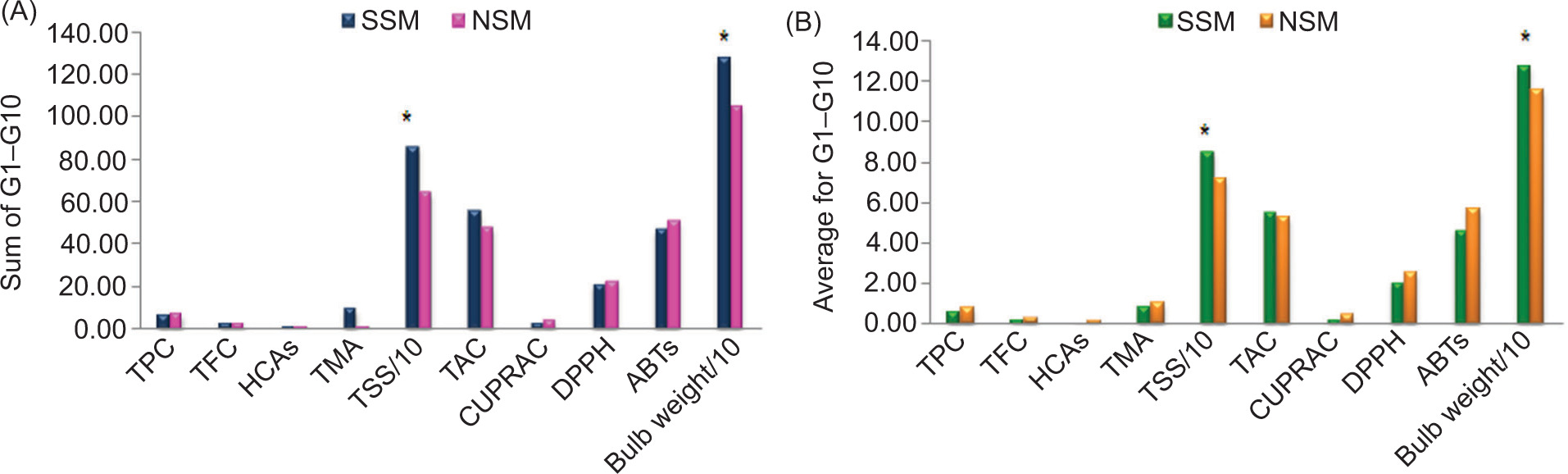
Figure 4. Sum values (A) and average values (B) of studied parameters in genotypes G1–G10 under the standard sowing method (SSM) and the new sowing method (NSM); Asterisk (*) denotes parameters which showed statistically significant differences between sowing methods.
Although SSM sowing resulted in higher sum values (genotypes G1–G10) of TFC (2.6 mg/g TE), HCAs (1.05 mg/g CGAE), TMA (9.43 mg/100 g C3GE), TSS (854 mg/g Glc), TAC (55.27 mg/g AAE), and bulb weight (1277.73 g), statistically significant differences were observed only for TSS and bulb weight (Figure 4).
For average values of studied parameters, TSS (85.41 mg/g Glc) and bulb weight (127.72 g) were statistically significantly higher in samples (G1–G10) under SSM (p<0.05). On the other hand, average values of other studied parameters (TPC – 0.81 mg/g GAE, TFC – 0.28 mg/g TE, HCAs – 0.19 mg/g CGAE, TMA – 1.06 mg/100 g C3GE) and antioxidant activity (TAC– 5.53 mg/g AAE, CUPRAC – 0.47 mg/g AAE, DPPH• – 2.51 µmol/g TE, and ABTS•+ – 5.66 µmol/g TE) were observed to be higher in onion genotypes sowed under the NSM method but with no significant differences (p> 0.05) (Figure 4). The sum and average values of HCAs and TMA were calculated only for violet-colored samples.
Pearson correlation analysis
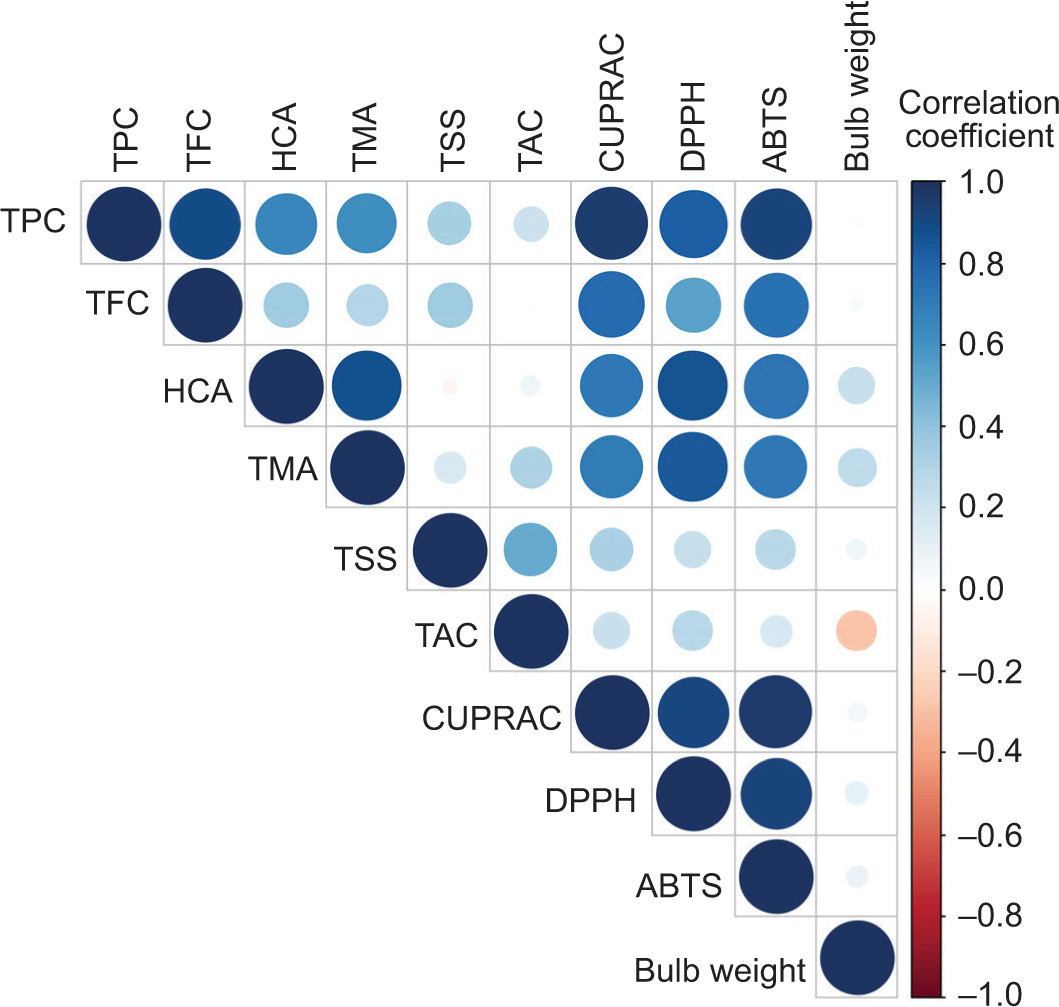
The Pearson correlation analysis presented the coefficients between various chemical parameters measured across 10 genotypes under two sowing methods (Figure 5).
Figure 5. Color correlation diagram for studied parameters in onion genotype samples.
For the purpose of this analysis, correlation coefficients (r) ≥ 0.70 were considered high, those between 0.30 and 0.70 were considered moderate, and those below 0.30 were considered weak or indicated no significant correlation.
A strong positive correlation was observed between TPC and TFC (r=0.896, p=0.000), suggesting that genotypes with higher TPC tend to also exhibit higher TFC. Similarly, TPC was statistically significantly correlated with HCAs (r=0.652, p=0.002), TMA (r=0.617, p=0.004), CUPRAC (r=0.950, p=0.000), DPPH• (r=0.823, p=0.000), and ABTS•+ (r=0.920, p=0.000), indicating that higher TPC was strongly associated with higher antioxidant capacities across various assays. However, TPC showed no significant correlation with TSS (r=0.332, p=0.153), TAC (r=0.212, p=0.370), or bulb weight (r=0.018, p=0.939), highlighting that TPC was not directly linked to TSS, TMA, or yield (as measured by bulb weight).
TFC also showed significant positive correlations with CUPRAC (r=0.782, p=0.000), DPPH• (r=0.542, p=0.014), and ABTS•+ (r=0.748, p=0.000), suggesting that TFC contributed substantially to antioxidant activity. No significant correlations were observed between TFC and HCA (r=0.359, p=0.121), TMA (r=0.297, p=0.204), TSS (r=0.358, p=0.121), TAC (r=0.015, p=0.949), or bulb weight (r=0.034, p=0.887), implying that the influence of flavonoids was more closely related to antioxidant capacity than other chemical parameters.
HCAs were strongly correlated with TMA (r=0.877, p=0.000), CUPRAC (r= 0.714, p=0.000), DPPH• (r=0.863, p=0.000), and ABTS•+ (r=0.735, p=0.000), indicating a close relationship between HCAs and TMA, as well as with antioxidant potential. However, HCAs did not show significant correlations with TSS (r=-0.038, p=0.873), TAC (r=0.076, p=0.751), or bulb weight (r=0.231, p=0.327).
TMA was positively correlated with CUPRAC (r=0.705, p=0.001), DPPH• (r=0.843, p=0.000), and ABTS•+ (r=0.715, p=0.000), reinforcing the association between TMA and antioxidant activity. No significant relationships were observed between TMA and TSS (r=0.169, p=0.476), TAC (r=0.312, p=0.180), or bulb weight (r=0.270, p=0.250). A moderate but significant correlation was observed between TSS and TAC (r=0.505, p=0.023).
Interestingly, CUPRAC, DPPH•, and ABTS•+ (three widely used assays to measure antioxidant capacity) are highly correlated with one another. CUPRAC shows robust correlations with DPPH• (r=0.919, p=0.000) and ABTS•+ (r=0.962, p=0.000), while DPPH• and ABTS•+ are similarly correlated (r=0.922, p=0.000), suggesting that these assays consistently reflect the antioxidant potential of the genotypes. Bulb weight does not show any significant correlation with antioxidant parameters such as CUPRAC (r=0.058, p=0.808), DPPH• (r=0.102, p=0.668), or ABTS•+ (r=0.085, p=0.722), reinforcing the idea that antioxidant content and yield were independent factors.
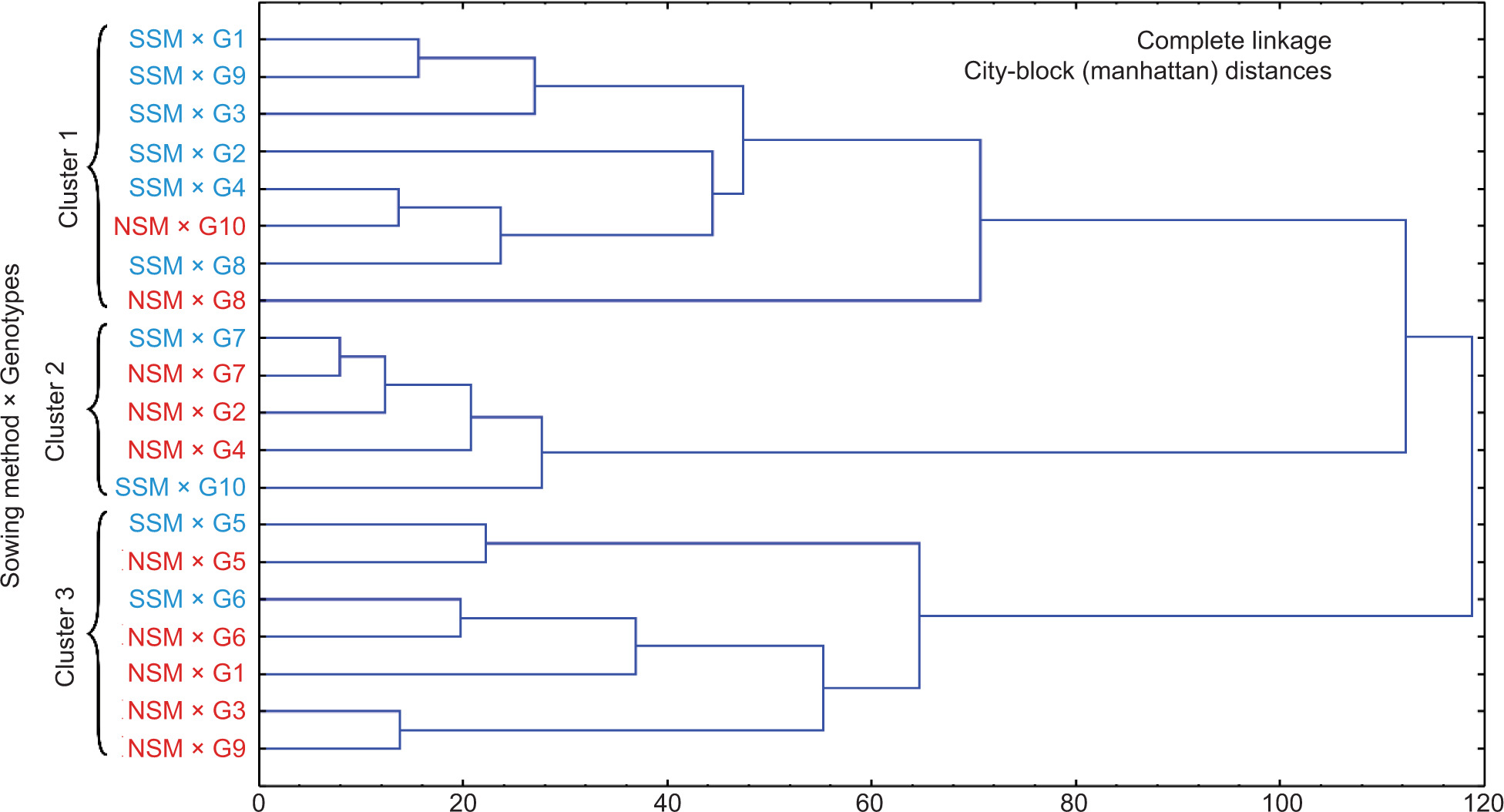
Cluster analysis
The CA of the treatments revealed three distinct groups, as shown in Figure 6. The first cluster included the treatments SSM × G1, SSM × G2, SSM × G3, SSM × G4, SSM × G8, SSM × G9, along with NSM × G8 and NSM × G10. These treatments were characterized by having the highest TAC and TSS. The second cluster comprised samples NSM × G2, NSM × G4, NSM × G7, as well as SSM × G7 and SSM × G10, all of which exhibited the highest bulb weight. Lastly, the third cluster included NSM × G1, NSM × G3, NSM × G5, NSM × G6, and NSM × G9, along with SSM × G5 and SSM × G6. These treatments were characterized by having the lowest content of bioactive compounds. The linkage distance between these clusters was approximately 120, indicating a relatively close relationship between the clusters.
Figure 6. Cluster analysis of studied parameters (TPC, TFC, HCA, TMA, TSS, TAC, CUPRAC, DPPH•, ABTS•+, and bulb weight) in onion genotypes.
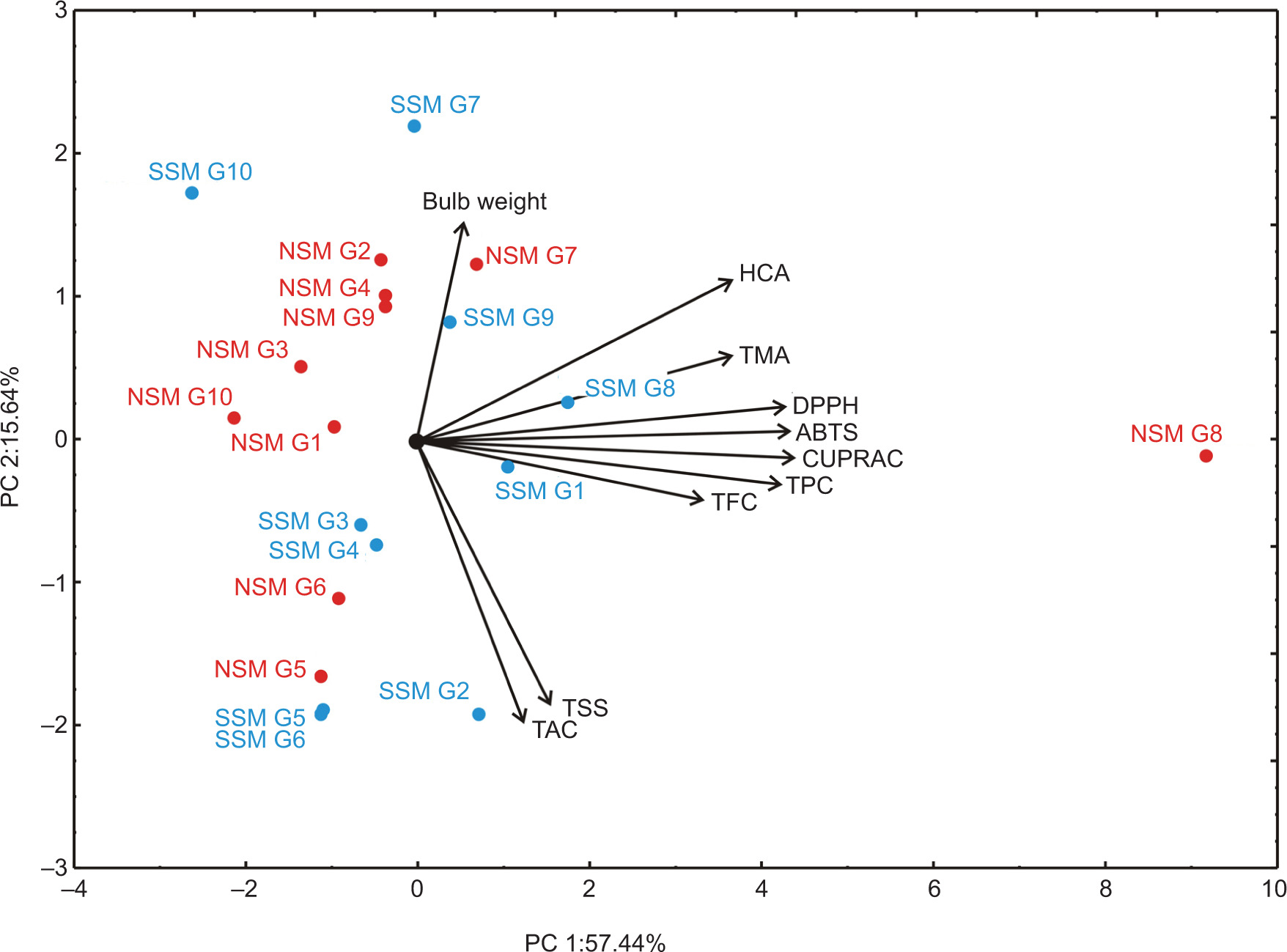
Principal component analysis
PCA was performed using the data presented in Figure 7. The PCA biplot, which explores the relationships among the parameters in onion samples across 10 genotypes under two distinct sowing methods, revealed that the first two principal components (PC1 and PC2) explained 73.08% of the total variance. This level of explained variance is considered adequate for reliable interpretation of the observed relationships. The PCA calculation showed the contribution of variables to the first two principal components, revealing the key factors that explain the variance in the dataset. For PC1, the variables with the highest contribution were CUPRAC (16.45% based on correlation), ABTS•+ (16.14%), and DPPH• (15.77%), indicating that these antioxidant-related parameters were the most influential in explaining the variation along this axis. TPC (15.38%) also had a substantial contribution, reflecting its strong association with PC1. TMA (11.56%) and HCAs (11.47%) provided moderate contributions. For PC2, the most significant contributors were TAC (–33.63%) and TSS (–29.15%), which had substantial negative contributions, indicating their inverse relationship with this component. On the positive side, bulb weight (20.40%) stands out as the most influential, with HCAs (10.88%) also showing a notable positive contribution. According to PCA analysis, PC1 reflects antioxidant activity, while PC2 highlights the role of bulb weight and certain bioactive compounds like TAC and TSS, offering two distinct dimensions of variance in the data.
Figure 7. PCA ordination of onion samples parameters (TPC, TFC, HCA, TMA, TSS, TAC, CUPRAC, DPPH•, ABTS•+, and bulb weight) based on their component correlations.
Discussion
Onion farmers aim for high yields while consumers value onion for its taste and secondary metabolites (preferably polyphenolic compounds), which are influenced by agronomic practices (Jeevitha et al., 2024; Ketema et al., 2018; Vojnović et al., 2024). Moreover, bulb color has become an important attribute in onions as consumers give importance to the positive attributes of vegetables, especially those rich in anthocyanins and flavonoids (Chalbi et al., 2023; Gorrepati et al., 2024; Zhang et al., 2016).
Results of the present study indicated that both the sowing method and genotype statistically significantly affected the content of the bioactive compound and bulb weight in onions (Table 2). For TFC, a key group of secondary metabolites, it was observed that the G1 × NSM treatment resulted in a statistically significantly lower TFC compared to G1 × SSM. This can be attributed to the higher plant density in SSM, which led to increased competition for water and nutrients, inducing stress and consequently promoting higher TFC synthesis. Changes in the sowing method can impact secondary metabolite composition, likely because of the competition for light, water, and nutrients (Björkman et al., 2011).
The content of flavonoids in onions is genetically determined. Moreover, the profile of flavonoids affects the color of onion. Among two main flavonoid classes found in onion, the anthocyanins attribute to red/purple color, whereas flavonols are responsible for the yellow and brown onion peel color (Rodrigues et al., 2017). In this experiment, genotype G8 exhibited the highest TFC, while G10 had the lowest, regardless of the sowing method (Table 2). A similar trend was found by Chalbi et al. (2023), who reported that red onions had the highest TFC (9.23 mg GAE g per dry weight (DW)), while white onions had the lowest (2.64 mg GAE g DW). Given the potent antioxidant, antimicrobial, and neuroprotective properties of TFC in onions (Ren and Zhou, 2021), genotype G8 could play a significant role in promoting human health as a fresh or cooked ingredient in diets worldwide.
Within the group of phenolic compounds, TFC plays a crucial role in defining the biological value of onions, with numerous health benefits, including antioxidant, anticancer, and antiviral properties (Ullah et al., 2020). TFC is also responsible for the pigmentation of flowers, which attracts pollinators. In addition, environments with flowers of various colors positively impact mental health (Zhang et al., 2023). In this study, the sowing method statistically significantly impacted the TFC in the tested genotypes. Specifically, the NSM treatment statistically significantly reduced TFC in G1, G3, and G4, likely because of the lower plant density in NSM compared to SSM.
Onion serves as an excellent example of a vegetable where genotype statistically significantly influences TFC (Marotti and Piccaglia, 2006). Phenolic content varied among different genotypes. In general, flavanol content is higher in yellow onions than in red onions (Gorrepati et al., 2024). Results of the present study, where G2 showed the highest and G10 the lowest TFC, are in line with literature. Likewise, Lee et al. (2005) found considerable variation in TFC across pepper (Capsicum annuum L.) genotypes. Yang et al. (2004) investigated onion genotypes of different colors, revealing that yellow onions (Western Yellow) had the highest TFC, while white onions (Western White) had the lowest, aligning with findings in this study; namely TFC was not detectable in the G10 sample with white skin.
HCAs are the largest class of phenolic acids that comprises a three-carbon side chain (C6–C3) structure. HCAs statistically significantly contribute to the flavor and nutritional quality of vegetables (Sova and Saso, 2020). As secondary metabolites, HCAs also play a role in plant defense against pathogens, UV radiation, and other environmental stresses. They provide health benefits for humans as well because of their potent antioxidant properties (Taofiq et al., 2017). Across both sowing methods (SSM and NSM), HCAs were measured only in genotypes with dark red (G7), violet (G8), and light red (G9) skin colors. Notably, HCAs were detected in dark bronze colored genotype G2 × NSM, but absent in the G2 × SSM treatment. However, a positive correlation was observed between HCAs and ABTS•+ assay results. This discrepancy may be because of the higher water availability in NSM (owing to lower plant density). This finding aligns with Park et al. (2023), who reported that water deficiency reduced HCA content in gomchwi (Ligularia fischeri (Ledeb.) Turcz.).
Onions vary in colors from dark red, yellow to white. Consumers are increasingly drawn to colorful onion varieties, with red and violet onions being especially preferred because of their visual appeal and perceived health benefits (Lee et al., 2015; Olsovska et al., 2024). These colors are mainly associated with anthocyanins and cationic flavylium ions (red), quinoidal bases (violet), and their colorless adducts (Frond et al., 2019). Anthocyanins play an essential role in human nutrition, primarily because of their strong antioxidant activity (Khoo et al., 2017). The presence of TMA was confirmed only in colored skin genotypes G7, G8 (SSM and NSM), and G9 (SSM). This corroborates with a study of Gorrepati et al. (2024) who reported that the mean total anthocyanin content was the highest in dark red genotypes, followed by its content in red, light bronze and white onion genotypes. Interestingly, a statistically significantly higher TMA was found in the G8 × NSM treatment, compared to G8 × SSM. This could be attributed to the lower plant density in NSM, leading to less shading and consequently increased light exposure, which promotes anthocyanin accumulation via the HY5 transcription factor in its biosynthesis pathway (Ma et al., 2021; Shin et al., 2013). This study confirmed the correlation between TMA content and antioxidant assays (CUPRAC, DPPH•, and ABTS•+). Similar results have been observed in onion, blackberry (Rubus fruticosus L.), and edible rose (family Rosaceae) (Ge et al., 2013; Lee et al., 2015; Marjanovic et al., 2021).
TSS is crucial in determining the texture, aroma, and flavor of onions. Moreover, the sugar content affects the storability of onion bulbs (Chope et al., 2007; Rutherford and Whittle, 1982). TSS values are generally associated with better storability, as they reduce water activity and lower the potential for microbial spoilage (Kiran et al., 2024; Sharma et al., 2020). In the present study, the sowing method and genotype statistically significantly altered the TSS content in some onion samples; for G2 × SSM, the TSS content was statistically significantly higher than in G2 × NSM. This could be explained by the higher competition for water in SSM, as TSS contributes to osmotic adjustment in plants experiencing water scarcity. For instance, in rice (Oryza sativa L. ssp. indica), drought conditions increased soluble sugar content in roots and leaves (Xu et al., 2015). However, the ability of osmoregulation is generally genotype-dependent (Munns and Tester, 2008). Similarly, TSS in the onion genotypes (G1, G3–G10) was not statistically significantly affected by the sowing method, supporting the finding by Chalbi et al. (2023) that TSS is primarily genotype-dependent. Higher TSS values are also associated with sweeter taste and improved flavor intensity (Patel et al. 2023; Wold et al., 2025). These attributes are highly valued by consumers, particularly in fresh market onions. Therefore, genotypes and sowing method that promote elevated TSS could have a significant impact on consumer satisfaction and market competitiveness.
Antioxidants in the diet are essential as they protect the human body from harmful free radicals (Luo et al., 2022; Zehiroglu et al., 2019). Compared to other vegetables like carrots (Daucus carota L.), celery (Apium graveolens L.), and tomatoes (Solanum lycopersicum L.), onions exhibit higher antioxidant activity (Kosewski et al., 2018), making them suitable for consumption in salads with fried and processed foods (e.g., grilled meats). In addition, onions are valuable in food preservation, as their antioxidant properties help prevent oxidative degradation when used as a spice (Tankoano et al., 2024). In this study, both genotype and sowing methods statistically significantly influenced antioxidant assay results (TAC, CUPRAC, DPPH•, and ABTS•+). Across both sowing methods, genotype G8 demonstrated the highest CUPRAC, DPPH•, and ABTS•+ activities. This might be due to its high TMA content, as a positive correlation between TMA and antioxidant activity was confirmed. Consequently, G8 is a strong candidate for breeding programs aimed at enhancing antioxidant properties. In addition, the NSM method often yields higher antioxidant activities in specific genotypes, especially in G8. The high antioxidant potential of specific onion genotypes has direct implications for consumer health, as regular intake of antioxidant-rich onions may help reduce the risk of chronic diseases associated with oxidative stress, including cardiovascular conditions and certain cancers (Desai et al., 2019; Tang et al., 2017). Furthermore, high-antioxidant onion genotypes, such as G8, hold significant market potential, meeting the growing consumer demand for functional foods rich in bioactive compounds. They are also valuable for both fresh consumption and use in the food processing industry.
This study confirmed a positive correlation between TFC and antioxidant activity (DPPH•, FRAP, and ABTS•+), consistent with Vojnović et al. (2023), in the Elenka F1 onion genotype and by Kim et al. (2024) in Korean onion genotypes. In addition, Ren and Zhou (2021) and Yang et al. (2004) both confirmed a strong correlation between TFC and antioxidant activity in onions. Chalbi et al. (2023) noted that antioxidant activity in onions, as measured by the ABTS•+ assay, depends on the genotype, which is in agreement with our findings. While this study established significant correlations between phytochemical content and antioxidant capacity, these relationships may reflect underlying causal mechanisms connected to genotype-specific metabolic pathways and responses to environmental stress, which warrants further investigation in future research.
During the experiment, certain deviations from long-term meteorological averages were observed, which could have influenced onion growth and phytochemical composition. Lower temperatures and reduced rainfall in March may have slowed early vegetative development and germination rates. Conversely, excessive rainfall in July and August likely prolonged the vegetative phase and delayed bulb maturation, as observed in the delayed harvest. In addition, higher temperatures and humidity in the summer months may have increased plant stress and stimulated the accumulation of secondary metabolites such as phenolic compounds and flavonoids, which are often associated with plant responses to abiotic stress. Similar effects of temperature and rainfall fluctuations on onion growth and secondary metabolite synthesis were reported by Caruso et al. (2014) and Bibi et al. (2022). These climatic variations highlight the importance of environmental conditions in shaping both bulb mass and quality parameters in onion production. These findings underline the importance of genotype–environment interactions, suggesting that breeding efforts should focus on selecting genotypes with stable phytochemical profiles and yield performance under variable climatic conditions.
Bulb mass is a vital yield component in onions. In this study, both genotype and sowing methods affected bulb mass (Table 1). Nourbakhsh and Cramer (2022) reported bulb masses ranging from 102.9 to 223.9 g, consistent with the results of the present study. In this study, genotype G10 had the highest bulb mass, with no significant difference between sowing methods. In G5, bulb mass was 19.71 g higher in NSM compared to SSM; however, this difference was not statistically significant This can be explained by the fact that increased growing space does not necessarily result in a significant increase in bulb formation; the genotypes appear to have reached their maximum genetic potential, regardless of the sowing method. In contrast, Caruso et al. (2014) found that sowing density statistically significantly affects bulb mass, likely because of differences in the genotypes used and because their onions were grown from seedlings rather than direct sowing. In terms of quantitative parameters, increasing plant density per hectare has been found to reduce root mass in Danshen (Salvia miltiorrhiza Bunge) and bulb mass in transplanted onions (Kanton et al., 2002; Xing et al., 2024).
Conclusions
This study demonstrates that both the sowing method and genotype play crucial roles in determining the phytochemical profile, yield, and overall quality of onions. Although bulb mass did not differ statistically significantly between the sowing methods for most genotypes, NSM offers clear economic advantages by reducing seed input per hectare. In addition, onions grown using NSM exhibited improved quality parameters, including higher antioxidant activity and elevated levels of key bioactive compounds, especially in red- and violet-skinned varieties, which are increasingly demanded by urban consumers. These results highlight NSM as a cost-effective alternative that not only sustains yield but also enhances crop quality. Future research should focus on integrating NSM with optimized fertilization strategies to further enhance yield, quality, and safety in onion production. In addition, testing these findings under different climatic conditions would help determine the stability and adaptability of both genotypes and sowing methods, ensuring their practical relevance in diverse environments.
Supplementary Materials
Table S1 is presented in the Supplementary material.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
Authors extend their sincere gratitude to the family farms Đukić and Salonski from Gospođinci and to the company Hoya V.S. d.o.o. from Subotica for technical and logistical support during the implementation of the experiment. Authors are also thankful to the students from the Faculty of Agriculture Novi Sad, course Field and Vegetable Crops for their assistance in the laboratory.
Author Contributions
Conceptualization was done by Đ.V. and J.P.; methodology was the concern of Đ.V. and Ž.I.; software was taken care of by L.P.; validation was done by Đ.V. and B.A.; formal analysis was carried out by S.K. and A.Ž.K.; investigation was done by S.K. and A.Ž.K.; resources were looked into by Đ.V.; data curation was done by L.P., B.A., and A.Ž.K.; writing—original draft preparation was the responsibility of Đ.V. and J.P.Dj.; writing—review and editing was taken care of by Đ.V. and J.P.Dj.; visualization and supervision were done by J.P.Dj. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Funding
This research was funded by the Ministry of Education, Science, and Technological Development of the Republic of Serbia, Grant Nos. 451-03-137/2025-03/ 200117, 451-03-4551/2024-04/17, and 451-03-137/2025-03/200116.
REFERENCES
Agricultural Extension Service Sombor. 2024. [cited 2024 September 09]. Available from: https://www.psssombor.rs/pis/
Alam, M.A., Rahman, M.A., Rahman, M.M., Hasan, M.M., Naher, S., Fahim, A.H.F., Mottalib, M.A., Roy, S., Islam, M.R., Mozumder, S.N., Alsuhaibani, A.M., and Hossain, A., 2023. Performance valuation of onion (Allium cepa L.) genotypes under different levels of salinity for the development of cultivars suitable for saline regions. Frontiers in Plant Science 14:1154051. 10.3389/fpls.2023.1154051
Alemu, D., Kitila, C., Garedew, W., Jule, L., Badassa, B., Nagaprasad, N., Seenivasan, V., Saka, A., and Ramaswamy, K., 2022. Growth, yield, and yield variables of onion (Allium cepa L.) varieties as influenced by plant spacing at DambiDollo, Western Ethiopia. Scientific Reports 12(1):20563. 10.1038/s41598-022-24993-x
Alkhateeb, O.A., Bardisi, A.A., Ismail, H., Ibrahem, S.K., Osman, A., Abou El-Yazied, A., Abdelaal, K., Alshammari, A., Rashed, A.A., Abdou, A.H., and Attaya, E., 2024. Response of onion fresh weight, nutrients uptake, nitrogen use efficiency, yield, and bulb quality to different nitrogen fertilization levels and plant density. Polish Journal of Environmental Studies 33(3):1–10. 10.15244/pjoes/176559
Aluta, U.P., Aderolu, A.Z., Ishola, I.O., Alyassin, M., Morris, G.A., and Olajide, O.A., 2023. Chemical characterisation of sulfated polysaccharides from the red seaweed Centroceras clavulatum and their in vitro immunostimulatory and antioxidant properties. Food hydrocolloids for health 3:100135. 10.1016/j.fhfh.2023.100135
Arena, D., Ben Ammar, H., Major, N., Kovačević, T.K., Goreta Ban, S., Al Achkar, N., Rizzo, G.F., and Branca, F., 2024. Diversity of the morphometric and biochemical traits of Allium cepa L. varieties. Plants 13:1727. 10.3390/plants13131727
Babbar, N., Oberoi, H.S., Sandhu, S.K., and Kumar Bhargav V., 2014. Influence of different solvents in extraction of phenolic compounds from vegetable residues and their evaluation as natural sources of antioxidants. Journal of Food Science and Technology 51: 2568–2575. 10.1007/s13197-012-0754-4
Bibi, N., Shah, M.H., Khan, N., Al-Hashimi, A., Elshikh, M.S., Iqbal, A., Ahmad, S., and Abbasi A.M., 2022. Variations in total phenolic, total flavonoid contents, and free radicals’ scavenging potential of onion varieties planted under diverse environmental conditions. Plants 11:950. 10.3390/plants11070950
Björkman, M., Klingen, I., Birch, A.N., Bones, A.M., Bruce, T.J., Johansen, T.J., Meadow, R., Mølmann, J., Seljåsen, R., Smart, L.E., and Stewart D., 2011. Phytochemicals of Brassicaceae in plant protection and human health–influences of climate, environment and agronomic practice. Phytochemistry 72(7):538–556. 10.1016/j.phytochem.2011.01.014
Caruso, G., Conti, S., Villari, G., Borelli, C., Melchionna, G., Minutolo, M., Russo, G., and Amalfitano C., 2014. Effects of transplanting time and plant density on yield, quality and antioxidant content of onion (Allium cepa L.) in southern Italy. Scientia Horticulturae 166:111–120 10.1016/j.scienta.2013.12.019
Chalbi, A., Chikh-Rouhou, H., Tlahig, S., Mallor, C., Garcés-Claver, A., Haddad, M., Sta-Baba, R., and Bel-Kadhi, M.S., 2023. Biochemical characterization of local onion genotypes (Allium cepa L.) in the arid regions of Tunisia. Polish Journal of Environmental Studies 32(1):15–26. 10.15244/pjoes/151867
Chaachouay, N., and Zidane, L., 2024. Plant-derived natural products: a source for drug discovery and development. Drugs and Drug Candidates 3(1):184–207. 10.3390/ddc3010011
Chope, A.G., Terry, A.L., and White, J.P., 2007. The effect of the transition between controlled atmosphere and regular atmosphere storage on bulbs of onion cultivars SS1, Carlos and Renate. Postharvest Biology and Technology 44(3):228–239. 10.1016/j.postharvbio.2006.12.018
Corrado, C.L., Micali, G., Mauceri, A., Bertin, S., Sunseri, F., Abenavoli, M.R., and Tiberini A., 2024. Study on Italian onion cultivars/ecotypes towards onion yellow dwarf virus (OYDV) infection. Horticulturae 10:73. 10.3390/horticulturae10010073
Degani, O., and Kalman, B., 2021. Assessment of commercial fungicides against onion (Allium cepa) basal rot disease caused by Fusarium oxysporum f. sp. cepae and Fusarium acutatum. Journal of Fungi 7:235. 10.3390/jof7030235
DuBois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., and Smith F., 1956 Colorimetric method for determination of sugars and related substances. Analytical Chemistry 28(3):350–356. 10.1021/ac60111a017
Frond, A.D., Iuhas, C.I., Stirbu, I., Leopold, L., Socaci, S., Andreea, S., Ayvaz, H., Andreea, S., Mihai, S., Diaconeasa, Z., and Carmen, S. 2019. Phytochemical characterization of five edible purple-reddish vegetables: anthocyanins, flavonoids, and phenolic acid derivatives. Molecules 24:1536. 10.3390/molecules24081536
Ge, Q., and Ma, X., 2013. Composition and antioxidant activity of anthocyanins isolated from Yunnan edible rose (An ning). Food Science and Human Wellness 2:68–74. 10.1016/j.fshw.2013.04.001
Gebretsadik, K., and Dechassa, N., 2018. Response of onion (Allium cepa L.) to nitrogen fertilizer rates and spacing under rain fed condition at Tahtay Koraro, Ethiopia. Scientific Reports 8: 9495. 10.1038/s41598-018-27762-x
Geries, M.S.L., El-Shahawy, A.T., and Moursi, A.E., 2021. Cut-off irrigation as an effective tool to increase water-use efficiency, enhance productivity, quality and storability of some onion cultivars. Agricultural Water Management 244:106589. 10.1016/j.agwat.2020.106589
Gökçe, A.F., Gökçe, Z.N.O., Junaid, M.D., and Chaudhry, U.K., 2023. Evaluation of biochemical and molecular response of onion breeding lines to drought and salt stresses. Scientia Horticulturae 311:111802. 10.1016/j.scienta.2022.111802
Golubkina, N., Amalfitano, C., Sekara, A., Tallarita, A., Pokluda, R., Stoleru, V., Cuciniello, A., Agafonov, F.A., Kalisz, A., Brînduşa Hamburdă,, S., and Caruso G., 2022. Yield and bulb quality of storage onion cultivars as affected by farming system and nitrogen dose. Scientia Horticulturae 293:110751. 10.1016/j.scienta.2021.110751
Gorrepati, K., Kumar, A., Ahammed Shabeer, T.P., Khan, Z., Satpute, P., Anandhan, S., Arunachalam, T., Yalamalle, V.R., Mahajan, V., and Singh, M., 2024. Characterization and evaluation of antioxidant potential of onion peel extract of eight differentially pigmented short-day onion (Allium cepa L.) varieties. Frontiers in Sustainable Food Systems 8:1469635. 10.3389/fsufs.2024.1469635
Ivanz, A., Boy, H., Rutillaa, A.J.H., Santos, K.A., Ty, A.M.T., Yu, A.I., Mahboob, T., Tangpoong, J., and Nissapatorn, V., 2018. Recommended medicinal plants as source of natural products: a review. Digital Chinese Medicine 1:131–142. 10.1016/S2589-3777(19)30018-7
Jeevitha, D., Chimmalagi, U., Adishesha, K., Hosamani, B.G., Vinay, V.T., and Dharmatti, R.P. 2024. Influence of planting methods on growth, yield and storage of onion (Allium cepa L.) var. Bhima Shakti. Indian Journal of Agricultural Research 58(5):885–888. 10.18805/IJARe.A-6217
Kanton, R.A.L., Abbey, L., Hilla, R.G., Tabil, M.A., and Jan, N.D., 2002. Density affects plant development and yield of bulb onion (Allium cepa L.) in Northern Ghana. Journal of Vegetable Crop Production 8(2):15–25. 10.1300/J068v08n0203
Ketema, S., Dessalegn, L., and Tesfaye B., 2018. Effect of planting methods on growth of onion (Allium cepa var. Cepa). Advances in Applied Physiology 3(1):8–13. 10.11648/j.aap.20180301.12
Kim, Y., Kim, Y.J., and Shin, Y., 2024. Comparative analysis of polyphenol content and antioxidant activity of different parts of five onion cultivars harvested in Korea. Antioxidants 13:197. 10.3390/antiox13020197
Khoo, H.E., Azlan, A., Tang, S.T., and Lim, S.M., 2017. Anthocyanidins and anthocyanins: colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food & Nutrition Research 61:1361779. 10.1080/16546628.2017.1361779
Kosewski, G., Górna, I., Bolesławska, I., Kowalówka, M., Więckowska, B., Główka, A.K., Morawska, A., Jakubowski, K., Dobrzyńska, M., Miszczuk, P., and Przysławski, J., 2018. Comparison of antioxidative properties of raw vegetables and thermally processed ones using the conventional and sous-vide methods. Food Chemistry 240:1092–1096. 10.1016/j.foodchem.2017.08.048
Kostić, A.Ž., Milinčić, D.D., Nedić, N., Gašić, U.M., Špirović Trifunović, B., Vojt, D., Tešić, Ž.L., and Pešić, M.B., 2021. Phytochemical profile and antioxidant properties of bee-collected artichoke (Cynara scolymus) pollen. Antioxidants 10:1091. 10.3390/antiox10071091
Kostić, A.Ž., Milinčić, D.D., Špirović Trifunović, B., Nedić, N., Gašić, U.M., Tešić, Ž.L., Stanojević, S.P., and Pešić, M.B., 2023. Monofloral corn poppy bee-collected pollen—a detailed insight into its phytochemical composition and antioxidant properties. Antioxidants 12:1424. 10.3390/antiox12071424
Kuralkara, P., and Kuralkar, S.V., 2021. Role of herbal products in animal production–an updated review. Journal of Ethnopharmacology. 278: 114246. 10.1016/j.jep.2021.114246
Lee, J.J., Crosby, M.K., Pike, M.L., Yoo, S.K., and Leskovar I.D., 2005. Impact of genetic and environmental variation on development of flavonoids and carotenoids in pepper (Capsicum spp.). Scientia Horticulturae 106(3):341–352. 10.1016/j.scienta.2005.04.008
Lee, E.J., Patil, B.S., and Yoo, K.S. 2015. Antioxidants of 15 onions with white, yellow and red colors and their relationship with pungency, anthocyanin, and quercetin. LWT–Food Science and Technology 63:108–114. 10.1016/j.lwt.2015.03.028
Luo, M., Zhou, L., Huang, Z., Li, B., Nice, E.C., Xu, J., and Huang, C., 2022. Antioxidant therapy in cancer: rationale and progress. Antioxidants 11:1128. 10.3390/antiox11061128
Ma, Y., Ma, X., Gao, X., Wu, W., and Zhou, B., 2021. Light induced regulation pathway of anthocyanin biosynthesis in plants. International Journal of Molecular Sciences 22:11116. 10.3390/ijms222011116
Marjanovic, A., Djedjibegovic, J., Lugusic, A., Sober, M., and Saso, L., 2021. Multivariate analysis of polyphenolic content and in vitro antioxidant capacity of wild and cultivated berries from Bosnia and Herzegovina. Scientific Reports 11:19259. 10.1038/s41598-021-98896-8
Marotti, M., and Piccaglia, R., 2006. Characterization of flavonoids in different cultivars of onion (Allium cepa L.). Journal of Food Science 67(3):1229–1232. 10.1111/j.1365-2621.2002.tb09482.x
Metrani, R., Singh, J., Acharya, P., Jayaprakasha, G.K., and Patil, B.S., 2020. Comparative metabolomics profiling of polyphenols, nutrients and antioxidant activities of two red onion (Allium cepa L.) cultivars. Plants 9:1077. 10.3390/plants9091077
Munns, R., and Tester, M., 2008. Mechanisms of salinity tolerance. Annual Review of Plant Biology 59:651–681. 10.1146/annurev.arplant.59.032607.092911
Nault, A.B., Hsu, L.C., and Hoepting A.C., 2012. Consequences of co-applying insecticides and fungicides for managing Thrips tabaci (Thysanoptera: Thripidae) on onion. Pest Management Science 69:841–849. 10.1002/ps.3444
Nourbakhsh, S.S., and Cramer, C.S., 2022. Onion plant size measurements as predictors for onion bulb size. Horticulturae 8:682. 10.3390/horticulturae8080682
Olsovska, K., Golisova, A., and Sytar, O. 2024. Optimizing nitrogen nutrient management for the sustainable enhancement of secondary metabolites and yield in onion cultivation. Sustainability 16(11):4396. 10.3390/su16114396
Park, Y.J., Kwon, D.Y., Koo, S.Y., Truong, T.Q., Hong, S.C., Choi, J., Moon, J., and Kim, S.M., 2023. Identification of drought-responsive phenolic compounds and their biosynthetic regulation under drought stress in Ligularia fischeri. Frontiers in Plant Science 14:1140509. 10.3389/fpls.2023.1140509
Piri, H., and Naserin, A., 2020. Effect of different levels of water, applied nitrogen and irrigation methods on yield, yield components and IWUE of onion. Scientia Horticulturae 268:109361. 10.1016/j.scienta.2020.109361
Rafie, R.M., Khoshgoftarmanesh, R.M., Shariatmadari, H., Darabi, A., and Dalir, N., 2017. Influence of foliar-applied zinc in the form of mineral and complexed with amino acids on yield and nutritional quality of onion under field conditions. Scientia Horticulturae 216:160–168. 10.1016/j.scienta.2017.01.014
Ren, F., and Zhou, S., 2021. Phenolic components and health beneficial properties of onions. Agriculture. 11:872. 10.3390/agriculture11090872
Rockenbach, I.I., Jungfer, E., Ritter, C., Santiago-Schübel, B., Thiele, B., Fett, R., and Galensa, R., 2012. Characterization of flavan-3-ols in seeds of grape pomace by CE, HPLC-DAD-MSn and LC-ESI-FTICR-MS. Food Research International 48(2):848–855. 10.1016/j.foodres.2012.07.001
Rodrigues, A.S., Almeida, D.P.F., Simal-Gándara, J., and Pérez-Gregorio, M.R., 2017. Onions: a source of flavonoids. In: Flavonoids–From Biosynthesis to Human Health. IntechOpen. 10.5772/intechopen.69896
Rutherford, P.P., and Whittle, R., 1982. The carbohydrate composition of onions during long term cold storage. Journal of Horticultural Sciences 57(3):349–356. 10.1080/00221589.1982.11515063
Sagar, N.A., Pareek, S., Benkeblia, N., and Xiao, J., 2022. Onion (Allium cepa L.) bioactives: chemistry, pharmacotherapeutic functions, and industrial applications. Food Frontiers 3:380–412. 10.1002/fft2.135
Schwinn, K.E., Ngo, H., Kenel, F., Brummell, D.A., Albert, N.W., McCallum, J.A., Pither-Joyce, M., Crowhurst, R.N., Eady, C., and Davies K.M., 2016. The onion (Allium cepa L.) R2R3-MYB gene MYB1 regulates anthocyanin biosynthesis. Frontiers in Plant Science 7:1865. 10.3389/fpls.2016.01865
Shin, D.H., Choi, M., Kim, K., Bang, G., Cho, M., Choi, S.B., Choi, G., and Park, Y.I., 2013. HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis. FEBS Letters 587(10):1543–1547. 10.1016/j.febslet.2013.03.037
Sova M., and Saso, L., 2020. Natural sources, pharmacokinetics, biological activities and health benefits of hydroxycinnamic acids and their metabolites. Nutrients 12(8):2190. 10.3390/nu12082190
Spence, C., 2015. On the psychological impact of food colour. Flavour 4:21. 10.1186/s13411-015-0031-3
Taofiq, O., González-Paramás, A.M., Barreiro, M.F., and Ferreira, I.C.F.R., 2017. Hydroxycinnamic acids and their derivatives: cosmeceutical significance, challenges and future perspectives, a review. Molecules. 22:281. 10.3390/molecules22020281
Teshika J.D., Zakariyyah A.M., Zaynab T., Zengin G., Rengasamy K.R.R., Pandian S.K., and Fawzi, M.M., 2019. Traditional and modern uses of onion bulb (Allium cepa L.): a systematic review. Critical Reviews in Food Science and Nutrition 59:S39–S70. 10.1080/10408398.2018.1499074
Ullah, A., Munir, S., Badshah, S.L., Khan, N., Ghani, L., Poulson, B.G., Emwas, A.H., and Jaremko, M., 2020. Important flavonoids and their role as a therapeutic agent. Molecules 25:5243. 10.3390/molecules25225243
Vojnović Đ., Maksimović, I., Tepić Horecki, A., Karadžić Banjac, M., Kovačević, S., Daničić, T., Podunavac-Kuzmanović, S., and Ilin Ž., 2023. Onion (Allium cepa L.) yield and quality depending on biostimulants and nitrogen fertilization—a chemometric perspective. Processes 11:684. 10.3390/pr11030684
Vojnović, Đ., Maksimović, I., Tepić Horecki, A., Milić, A., Šumić, Z., Žunić, D., Adamović, B., and Ilin, Ž., 2024. Biostimulants improve bulb yield, concomitantly affecting the total phenolics, flavonoids, and antioxidant capacity of onion (Allium cepa). Horticulturae 10:391. 10.3390/horticulturae10040391
Vuković, S., Moravčević, Đ., Gvozdanović-Varga, J., Kostić, A.Ž., Vujošević, A., Kilibarda, S., and Pećinar, I., 2022. Raman spectroscopy as a useful tool for tentative identification of nutritional ingredients and distinction of allium species. Biology and Life Sciences Forum 16(1):21. 10.3390/IECHo2022-12480
Vuković, S., Popović-Djordjević, J.B., Kostić, A.Ž., Pantelić, N.D., Srećković, N., Akram, M., Laila, U., and Katanić Stanković, J.S., 2023a. Allium species in the Balkan Region—major metabolites, antioxidant and antimicrobial properties. Horticulturae 9:408. 10.3390/horticulturae9030408
Vuković, S., Moravčević, D., Gvozdanović-Varga, J., Dojčinović, B., Vujošević, A., Pećinar, I., Kilibarda, S., and Kostić, A.Ž., 2023b. Elemental profile, general phytochemical composition and bioaccumulation abilities of selected Allium species biofortified with selenium under open field conditions. Plants 12:349. 10.3390/plants12020349
Xing Z., Bi G., Li T., Zhang Q., and Knight P.R., 2024. Effect of plant density on growth and bioactive compounds in Salvia miltiorrhiza. Agronomy 14:1157. 10.3390/agronomy14061157
Xu, W., Cui, K., Xu, N.L., Huang, J., and Peng, S., 2015. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiologiae Plantarum. 37:9. 10.1007/s11738-014-1760-0
Yang, J., Meyers, K.J., Van der Heide, J., and Liu, R.H., 2004. Varietal differences in phenolic content and antioxidant and antiproliferative activities of onions. Journal of Agricultural and Food Chemistry 52:6787–6793. 10.1021/jf0307144
Zehiroglu, C., and Ozturk Sarikaya, S.B., 2019. The importance of antioxidants and their place in today’s scientific and technological studies. Journal of Food Science and Technology 56(11):4757–4774. 10.1007/s13197-019-03952-x
Zhang, L., Dempsey, N., and Cameron, R., 2024. ‘Blossom Buddies’—how do flower colour combinations affect emotional response and influence therapeutic landscape design? Landscape and Urban Planning 248:105099. 10.1016/j.landurbplan.2024.105099
Zhang, S.L., Deng, P., Xu, Y.C., Lü, S.W., and Wang J.J., 2016. Quantification and analysis of anthocyanin and flavonoid compositions, and antioxidant activities in onions with three different colors. Journal of Integrative Agriculture 15(9):2175–2181. 10.1016/S2095-3119(16)61385-0
Zhao, X.X., Lin, F.J., Li, H., Li, H.B., Wu, D.T., Geng, F., Ma, W., Wang, Y., Miao, B.H., and Gan R.Y., 2021. Recent advances in bioactive compounds, health functions, and safety concerns of onion (Allium cepa L.). Frontiers in Nutrition 8:669805. 10.3389/fnut.2021.669805
Supplementary
Table S1. Applied pesticides during the onion experiment.
| Commercial name | Active ingredient | Application rate | Target pest/disease/weed |
|---|---|---|---|
| Dicarzol® (Belchim Crop Protection, Belgium) (i) | Formetanate hydrochloride 582 g/kg | 0.5 L/ha | Thrips tabaci Lindeman |
| Exirel® (FMC International, Switzerland) (i) | Cyantraniliprole 100 g/L | 0.5 L/ha | Delia antique Meigen, Thrips spp. |
| Signum® (BASF SE, Germany) (f) | Boscalid 267 g/kg + pyraclostrobin 67 g/kg | 0.7–1 kg/ha | Peronospora destructor (Berk) Casp. |
| Infinito (Bayer, Germany) (f) | Propamocarb hydrochloride 625 g/L + fluopicolide 62.5 g/L | 1.6 L/ha | Peronospora destructor (Berk) Casp. |
| Fusilade Forte® (Syngenta Crop Protection, Switzerland) (h) | Fluazifop-p-butyl 150 g/L | 0.8 L/ha | Sorghum halepense (L.) Pers, Agropyron repens (L.) P. Beauv. |
| Lentagran® (Belchim Crop Protection, Belgium) (h) | Pyridate 450 g/kg | 2.5 kg/ha | Amaranthus retroflexus L. , Abutilon theophrasti Medik. |
| Stomp®Aqua (BASF SE, Germany) (h) | Pendimethalin 455 g/L | 2 L/ha | Capsella bursa-pastoris (L.) Medik. , Chenopodium album L ., Amaranthus retroflexus L. |
| Inex® (Cosmocel, Mexico) (a) | Ethoxylated fatty alcohols 20.2%, polydimethylsiloxane 1% | 2.5 L/ha | / |
i–Insecticides; f–Fungicides; h–Herbicides; a–Adjuvants.